Доклад про кожу человека 4 класс: Мы и окружающий мир. 4-й класс. «Кожа, особенности кожного покрова, функции кожи» (урок по системе Л.В. Занкова)
Анализы на ВПЧ цена — сдать анализ на папилломавирус человека в СПб
Вирус папилломы человека (ВПЧ) — одна из самых распространённых инфекций, передающихся половым путём и при контакте «кожа-к-коже». Нет точной статистики, сколько людей в мире заражены ВПЧ, потому что у большинства заражённых вирус никак себя не проявляет. Биологи считают, что около 80 % населения земли — носители ВПЧ.
Опасность вируса в том, что некоторые его типы могут привести к онкологии. 98 % рака шейки матки (по данным учёных США — 100 %) связаны с ВПЧ. Вирус провоцирует рак влагалища, вульвы, анального канала, мужских половых органов, горла, рта. Причём смертельно опасное заболевание может развиться спустя годы, десятилетия после заражения, если возникнут благоприятные для этого условия.
Всего в группу ВПЧ входит более 170 вирусов и штаммов, из них примерно 40 передаются половым путём, а 13 способны вызвать рак.
Заражение отдельными типами ВПЧ — при наличии высокого иммунитета — проходит незаметно, бесследно для организма. Другие штаммы заставляют клетки кожи усиленно делиться, в итоге появляются бородавки, папилломы, кондиломы («венерические бородавки»). Третьи типы, интегрируя в ДНК человека, действуют как онкогены и способствуют злокачественной трансформации клеток, росту опухолей.
Это важно! ВПЧ не является достаточным фактором для развития онкологического заболевания. Но он — один из важнейших онкофаторов, «провокатор» озлокачествления клеток.
Чтобы началось перерождение здоровой ткани в раковую, необходимо сочетание нескольких условий, в их числе — сбои в работе иммунной системы. Именно иммунитет — главный защитник в том числе и от увеличения вирусной нагрузки ВПЧ.
Очень важно периодически сдавать анализа на ВПЧ, чтобы предотвратить опасность, вызванную вирусом. Так, можно вылечить предраковое состояние шейки матки.
ВПЧ: способы заражения, симптомы, онкогенные типы вируса
Основные способы заражения:
-
вагинальный, оральный, анальный секс с человеком, заражённым ВПЧ;
-
при контактах «кожа-к-коже» с человеком, заражённым папилломавирусом, или контакте с поверхностями, которых касался заражённый человек — бытовое заражение возможно, если на коже имеются порезы, ссадины, другие повреждения;
-
при родах — от матери к ребёнку.
Последние исследования американских и европейских учёных показали, что велика вероятность заражения ВПЧ в медицинских учреждениях — при переливании крови, использовании медицинского оборудования, вдыхании вирусных частиц, например — при лазерной абляции или электрокоагуляции кондилом.
Сложность борьбы с неполовыми формами заражениями в том, что вирус чрезвычайно устойчив к большинству дезинфицирующих средств. ВПЧ — первый вирус, оказавшийся нечувствительным к инактивации (обработке) глутаровым альдегидом (средством для стерилизации хирургических инструментов, требующих абсолютной чистоты). Перед врачами и технологами встала проблема обеззараживания приборов, которые нельзя автоклавировать и подвергать воздействию агрессивных химических соединений.
Симптомы и типы вируса папилломы человекаСимптомы заражения различаются в зависимости от типа ВПЧ. Некоторые типы, например — ВПЧ5, сохраняются в организме человека без клинических симптомов и могут быть обнаружены только специальными исследованиями. Штаммы ВПЧ1, 2, 4, 7, 22, 63 вызывают образование бородавок на руках, ногах, подошвах.
Типы 6, 11, 42, 44 способны вызвать развитие генитальных бородавок, папилломатоза гортани; 6, 16, 18, 31 и другие — анальной дисплазии; 60 — вирусной кисты.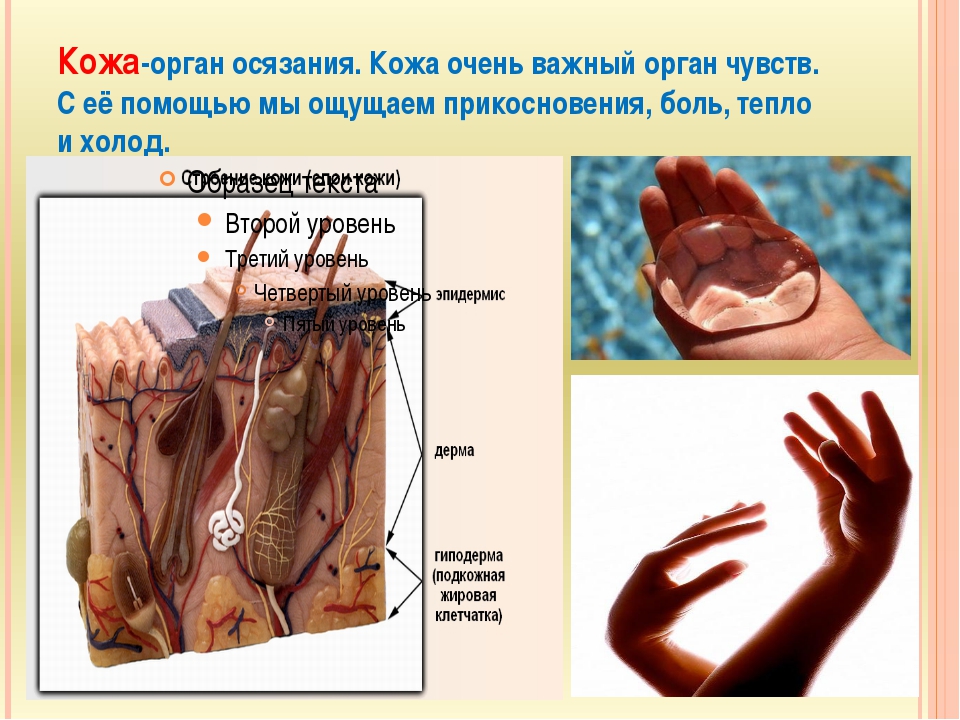
К раку половых органов способны привести штаммы 26, 53, 66. Штаммы с высоким онкориском — 33, 35, 39, 51, 52, 56, 58, 59. Самым высоким риском трансформации клеток в злокачественные обладают типы 16, 18, 31, 45.
Когда назначаются
Анализы на ВПЧ нужно пройти каждому взрослому человеку самостоятельно, без назначения, т. к. вероятность, что вы заражены — 8 из 10.
Направление на анализ обычно выдаётся дерматологом, урологом, гинекологом при наличии характерных внешних признаков или заболеваний, причиной которых может стать вирус.
Анализы на ВПЧ сдают при планировании беременности, при выявлении причин и лечении бесплодия, патологий беременности и вынашивания. В этом случае анализы сдают оба партнёра.
Факторами риска и поводом для сдачи анализа у женщин также являются:
-
ранняя половая жизнь;
-
отношения с разными, иногда сразу несколькими половыми партнёрами;
-
общие хронические, гинекологические заболевания, патологии;
-
слабый иммунитет.


Факторами риска и поводом для сдачи анализа у мужчин также являются:
-
множественные половые контакты;
- половые контакты с женщинами, заражёнными ВПЧ;
-
плохая гигиена;
-
сужение крайней плоти;
-
слабый иммунитет.
Кольпоскопическое исследование
Кольпоскопия — осмотр с помощью кольпоскопа влагалищной части шейки матки, входа и стенок влагалища. Это простой, недорогой, но высокоинформативный метод диагностирования заболеваний шейки матки.
Клиническое значение имеет расширенная кольпоскопия с применением нескольких тестов — с 3-процентной уксусной кислотой, йодным раствором Люголя. Тесты выявляют различные типы эпителия, позволяют оценить размеры и качество патологических образований (при их наличии), сосудистый рисунок, качество шеечных желёз.
Тесты выявляют различные типы эпителия, позволяют оценить размеры и качество патологических образований (при их наличии), сосудистый рисунок, качество шеечных желёз.
Во время кольпоскопии проводят прицельную биопсию с наиболее атипично изменённых участков.
Цитологическое исследование
Задача цитологического исследования шеечных мазков (тест Папаниколау, пап-тест) — выявление специфических для ВПЧ-инфекции клеток — койлоцитов и дискератоцитов.
Подтверждением папилломовирусной инфекции считается обнаружение койлоцитов, трансэпителиальной лимфоцитарной инфильтрации, базально-клеточной гиперплазии в биоптате (биоматериале, взятом на исследование).
Пап-тест обязателен для:
-
женщин после 30 лет;
-
женщин, у кого был ранее диагностирован ВПЧ;
-
женщин, у кого во время кольпоскопии обнаружили зоны с изменённым эпителием.

По результатам пап-теста определяют класс опасности для здоровья женщины: 1—2 класс — без подозрения на рак, 3 класс — подозрение на онкологию, 4—5 класс — наличие раковых клеток в малом или большом количестве.
К недостаткам цитологического исследования относят сложность исполнения, высокие квалификационные требования к врачу-цитологу. Потому проходить исследование нужно в диагностических центрах и лабораториях, персонал которых постоянно подтверждает свой профессионализм.
Гистологическое исследование
Гистологический метод обнаружения ВПЧ можно было бы считать золотым стандартом диагностики вируса, однако мешает его высокая стоимость, невозможность частого проведения и не всегда точный прицельный забор биоптата из шейки матки. Для проведения гистологической диагностики также требуются специалисты очень высокой квалификации.
Поэтому гистологическое исследование биоптата часто служит дополнением к пап-анализу.
ПЦР диагностика папилломавируса
Полимеразная цепная реакция (ПЦР) относится к высокоразрешающим технологиям детекции нуклеиновых кислот. Современные ПЦР тест-системы обладают высокой чувствительностью, используются не только для выявления ВПЧ, но и вирусной нагрузки на организм (количественный показатель заражённости) главных клинически значимых генотипов (16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59), которые ответственны за почти 94 % случаев тяжёлых цервикальных дисплазий и рака шейки матки. ПЦР тест-системы используют также для обнаружения штаммов ВПЧ 6 и 11.
Такие возможности тест-системы позволяют прогнозировать течение ВПЧ-инфекции, оценивать эффективность терапии. Установлено, что папилломавирусная инфекция имеет дозозависимый эффект: чем выше концентрация ДНК вируса в исследуемом материале, тем выше риск развития неоплазии и раковой опухоли.
В результатах теста указывают концентрацию ВПЧ:
-
Lg < 3 — папилломавирус обнаружен в клинически незначимом количестве;
-
Lg 3–5 — папилломавирус обнаружен в клинически значимом количестве;
-
Lg > 5 — папилломавирус обнаружен в высокой концентрации, вирусная нагрузка на организм высокая.
Виды исследуемого материала, правила забора:
-
для пап-теста — шеечный мазок;
-
для цитологического исследования — биоптат, взятый прицельно с атипично изменённых участков слизистой.
-
для ПЦР-теста — соскоб клеток слизистых оболочек генитального тракта.
Мазок у женщин берут из цервикального канала, у мужчин — из уретры. Для мазка используют мягкую щётку или ватный тампон. Их бережно вводят в канал, затем осторожно вынимают, вращая. На поверхности щётки/тампона остаются эпителиальные клетки, нужные для исследования.
Для мазка используют мягкую щётку или ватный тампон. Их бережно вводят в канал, затем осторожно вынимают, вращая. На поверхности щётки/тампона остаются эпителиальные клетки, нужные для исследования.
Взятие биоматериала не проводится во время менструации, за 5 дней до её начала и в течение 5 дней по окончании. Нельзя проходить исследование, если есть воспалительные процессы.
За 2 суток до забора биоматериала женщинам и мужчинам нужно воздержаться от сексуальных контактов. Женщинам нельзя в течение 48 часов использовать вагинальные кремы, лекарства, суппозитории, спринцевания, тампоны, вместо ванны нужно принимать душ. Мазок берут до проведения любых гинекологических манипуляций или через 2 суток после них.
Если биоматериал берут из уретры, то от последнего мочеиспускания до забора биоматериала должно пройти не менее 90 минут.
При заборе биоптата для гистологического исследования соблюдаются те же правила, что и при подготовке к забору мазка. После биопсии в течение 2—3 недель нельзя:
После биопсии в течение 2—3 недель нельзя:
вступать в половые контакты,
-
испытывать значительные физические нагрузки,
-
перегреваться (баня, сауна, жаркая погода),
-
купаться в открытом водоёме или бассейне,
-
принимать препараты, разжижающие кровь;
-
использовать вагинальные средства.
Если возникнет кровотечение, следует пользоваться только прокладками (не тампонами!)
На точность результата могут влиять антибиотики, пробиотики, местные антисептики, которые вы принимали/применяли даже 2 месяца назад. Перед сбором материала нужно рассказать врачу обо всех лекарственных средствах, которыми вы пользовались или пользуетесь.
Стоимость исследования на заражение ВПЧ зависит от вида исследования, охвата штаммов вируса, определения типа/без определения, расчёта вирусной нагрузки.
Цены на виды исследований в медицинских подразделениях АО «СЗДЦМ» представляют собой разнообразные комбинации по охватности, подробности, прогностической ценности.
Какой способ исследования выбрать, вам подскажет врач — гинеколог, дерматолог, венеролог. Если вы сдаёте анализы по собственной инициативе, выберите исследование на наличие онкогенных штаммов ВПЧ.
Анализы на ВПЧ вы можете сдать в медицинских подразделениях АО «СЗДЦМ», расположенных в Санкт-Петербурге, Ленинградской области, в Великом Новгороде, Старой Руссе и других городах.
Чтобы найти ближайший к вам пункт, воспользуйтесь интерактивной картой или перечнем медицинских учреждений АО «СЗДЦМ».
Во всех наших отделениях — терминалах и центрах — вас встретят внимательные, опытные специалисты с высокой квалификацией. Медицинские учреждения АО «СЗДЦМ» оснащены современным оборудованием, лабораторными материалами, одноразовыми инструментами и расходными материалами.
Мы гарантируем вам точность исследований, заботливое отношение, полную конфиденциальность ваших личных данных и результатов обследования.
Будьте здоровы! А для этого вовремя и регулярно проходите важные обследования в АО «СЗДЦМ».
Мы поможем вам сохранить и вернуть здоровье!
Как одеваться в школу и сколько должен весить учебник – что говорят правоохранители Подмосковья
Перед началом учебного года прокуратура Московской области решила напомнить родителям учащихся и педагогам о правах детей на образование, а также разъяснить существующие требования к школьной форме и весу учебников, разместив на своем сайте соответствующие памятки. В связи с этим родители и педагоги должны обратить особое внимание на удовлетворение образовательных потребностей детей. Подробнее о том, какие права школьника необходимо соблюдать, какие требования к школьной форме существуют в Московской области и сколько должны весить школьные учебники и портфель, читайте в материале портала mosreg. ru.
ru.
Марина Захарова: «Подмосковье входит в пятерку лучших регионов страны по уровню образования» >>
Гарантированное право на образование и защита детей
Право граждан на образование закреплено Конституцией Российской Федерации.
Как сообщается на сайте прокуратуры Московской области, право на образование гарантируется гражданину независимо от пола, расы, национальности, языка, происхождения, имущественного, социального и должностного положения, места жительства, отношения к религии, убеждений, принадлежности к общественным объединениям и др., согласно Федеральному закону «Об образовании в Российской Федерации».
Условия для обучения ребенка должны соответствовать особенностям психофизического развития и состояния здоровья, поэтому обучение может осуществляться по индивидуальному учебному плану.
При этом, согласно пункту 1 статьи 34 данного закона, ребенок может сам выбирать образовательную организацию и форму получения образования.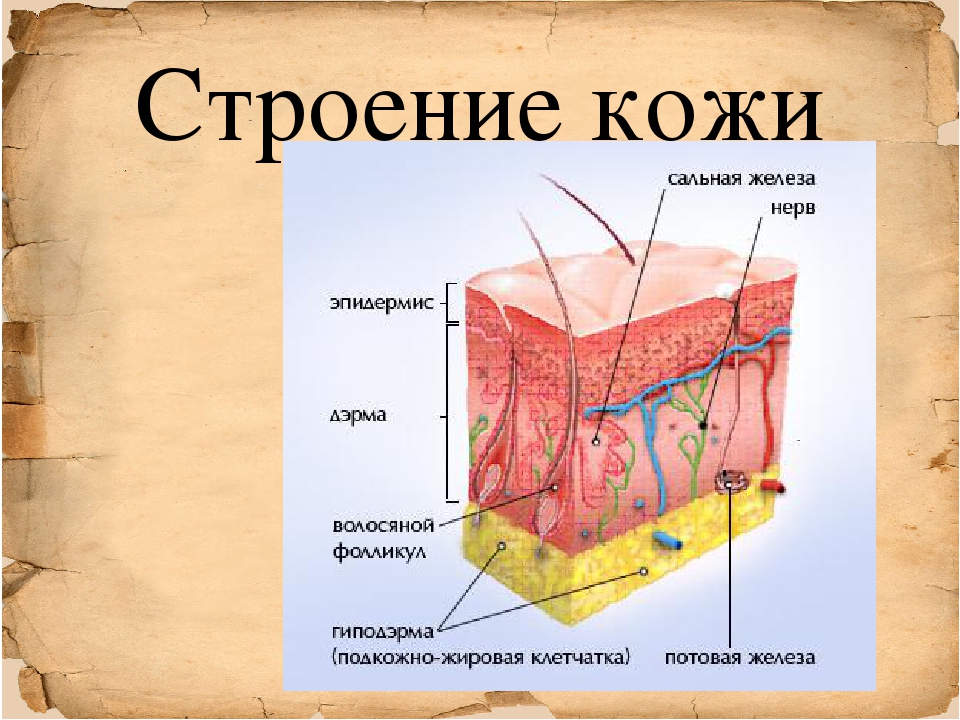
Родителям, опекунам, школьным педагогам и администрации необходимо строго соблюдать и защищать права детей в сфере осуществления образовательной деятельности. В школе ребенок должен быть защищен от всех форм физического и психического насилия, оскорбления личности. Он имеет право на охрану жизни и здоровья, свободу совести и др. Каждый обучающийся, попавший в трудную жизненную ситуацию, должен своевременно получить необходимую помощь, в том числе посредством работы школьной службы медиации.
Поскольку ни один случай нарушения прав детей при осуществлении образовательной деятельности не должен оставаться без внимания, органы прокуратуры проверяют любую поступающую информацию о нарушениях прав детей в сфере образования и принимают меры для восстановления законности и защиты их интересов.
Узнайте, как перевести ребенка в новую школу в Подмосковье>>
О запрете коррупции в школе
В сфере образования недопустимы любые проявления коррупции:
- незаконное взимание платы за зачисление, восстановление, перевод в государственные и муниципальные образовательные организации, проведение промежуточной и итоговой аттестации, выдачу документов об образовании,
- возмездное оказание обучающимся образовательных услуг, финансируемых за счет бюджетных средств,
- поборы, понуждение обучающихся и родителей к внесению благотворительных пожертвований на укрепление материальной базы школы, охрану зданий, приобретение учебников, учебных пособий, спортивного инвентаря и средств обучения, осуществление внеклассной работы, расходы на которую должны нести сами образовательные организации или их учредители.

Зачем нужна школьная форма единого образца
Школьная форма способствует тому, чтобы ученики чувствовали свою принадлежность к школе, настраивает на получение знаний. К тому же дети не должны ощущать различий в социальном и материальном положении, религиозной принадлежности.
«Статьей 38 Федерального закона от 29.12.2012 № 273-ФЗ «Об образовании в Российской Федерации» организациям, осуществляющим образовательную деятельность, предоставлено право устанавливать требования к одежде обучающихся (в том числе общему виду, цвету, фасону, знакам отличия) и правила ее ношения, которые закрепляются локальным нормативным актом соответствующей организации, принимаемым с учетом мнения совета обучающихся, совета родителей, а также представительного органа работников организации», — сообщается на сайте прокуратуры Московской области.
При этом Типовые требования к школьной форме установлены распоряжением Министерства образования Московской области от 26. 01.2015 № 1.
01.2015 № 1.
Во-первых, школьная форма должна соответствовать гигиеническим требованиям к детской одежде и контактирующим с кожей человека изделиям, требованиям безопасности продукции, предназначенной для детей и подростков, а также погоде и месту проведения учебных занятий, температуре помещения. Во-вторых, одежда должна соответствовать нормам делового стиля и носить светский характер.
На одежде школьников могут быть эмблемы, нашивки, значки и другие отличительные знаки образовательной организации (класса, параллели классов).
Что касается частных школ, то они могут не следовать типовым требованиям к фасону и цветовой гамме школьного гардероба. Тем не менее они должны соблюдать требования федерального законодательства, направленные на охрану жизни, здоровья и нормальное развитие несовершеннолетних при установлении собственной школьной формы.
Школьный портал Подмосковья: электронный дневник, домашнее задание и общение онлайн>>
Требования к повседневной и парадной форме
Мальчики и юноши в качестве повседневной одежды должны носить классические брюки, пиджак (жилет) нейтральных или неярких цветов (уместна ткань в клетку или полоску классических цветов), однотонную рубашку сочетающегося с ними цвета. Что касается аксессуаров, можно носить галстук, поясной ремень, в холодное время года — джемпер, свитер, пуловер нейтральных или неярких цветов (также допустима ткань в клетку или полоску).
Что касается аксессуаров, можно носить галстук, поясной ремень, в холодное время года — джемпер, свитер, пуловер нейтральных или неярких цветов (также допустима ткань в клетку или полоску).
Девочки и девушки в качестве повседневной одежды могут носить жакет, жилет, юбку, сарафан или брюки классического стиля нейтральных или неярких цветов (уместна ткань в классическую клетку или полоску). Блузка должна быть непрозрачной, длиной ниже талии, а платье можно дополнить белым или черным фартуком, воротником. Длина платьев, юбок и сарафанов должна варьироваться в пределах 10 см от верхней и нижней границы колена.
В дни проведения праздников и торжественных мероприятий школьная одежда дополняется светлой рубашкой для мальчиков и блузкой для девочек, праздничным аксессуаром.
На занятиях физкультурой и спортом должна использоваться спортивная форма, которая включает футболку, спортивные шорты, спортивные брюки, спортивный костюм. Что касается обуви, это кеды или кроссовки.
Узнайте, какие льготы могут получить школьники в Подмосковье>>
Что нельзя надевать в школу
Школьникам не рекомендуется носить одежду, обувь и аксессуары с острой фурнитурой, которая может травмировать.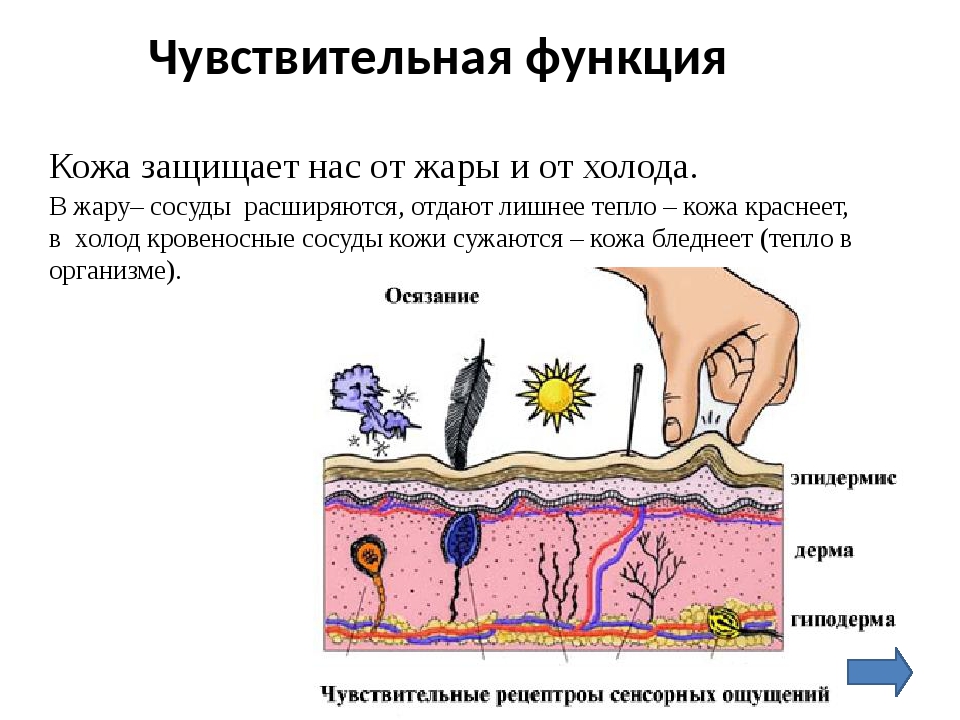 Также в школе нельзя носить одежду с яркими надписями и изображениями (кроме специальной спортивной одежды) или пропагандирующими психоактивные вещества и противоправные действия, а также с символикой асоциальных неформальных молодежных объединений.
Также в школе нельзя носить одежду с яркими надписями и изображениями (кроме специальной спортивной одежды) или пропагандирующими психоактивные вещества и противоправные действия, а также с символикой асоциальных неформальных молодежных объединений.
Запрещены атрибуты одежды, закрывающие лицо (кроме случаев, когда это необходимо по состоянию здоровья обучающихся).
Девочкам и девушкам нельзя носить в школу брюки и юбки с заниженной талией или высокими разрезами, декольтированные платья и блузки. Также запрещается носить в школу обувь на толстой платформе, туфли на каблуках выше 7 сантиметров и массивные украшения.
Как согласовать проведение спортивных соревнований в Подмосковье>>
Ограничения по весу для учебников и портфелей
Современным школьникам приходится носить в своем рюкзаке большое количество учебников и пособий. Ношение тяжестей и чрезмерная физическая нагрузка отрицательно сказывается на физическом и эмоциональном состоянии подростка, отражается на его работоспособности, а впоследствии может стать причиной заболеваний опорно-двигательного аппарата школьника.
В связи с этим законодательством Российской Федерации определены обязательные для издателей ограничения массы учебников, установленные пунктом 3.1.3 СанПиН 2.4.7.1166-02 «Гигиена детей и подростков. Гигиенические требования к изданиям учебным для общего и начального профессионального образования» от 20.11.2002 № 38.
Так, учебник младшего школьника (1–4 класс) должен весить не более 300 граммов (а издание, предназначенное только для работы в классе – не более 500 граммов), для 5–6-х классов – не более 400 граммов, для 7–9-х классов – не более 500 граммов, для 10–11-х классов – не более 600 граммов.
Учебники, вес которых более чем на 10% превышает установленные пределы, не допускаются к использованию.
Также существуют нормативы веса для портфелей, ранцев, рюкзаков и школьно-письменных принадлежностей, которые определены Техническим регламентом таможенного союза «О безопасности продукции, предназначенной для детей и подростков» от 23.09.2011 № 797.
Согласно регламенту, вес ученического ранца, сумки, портфеля или рюкзака для учащихся начальных (1–4-х) классов не может превышать 700 граммов, а для учащихся средних и старших (5–11-х) классов – 1000 граммов. При этом каждое изделие должно иметь маркировку с указанием возраста пользователей.
При этом каждое изделие должно иметь маркировку с указанием возраста пользователей.
Оптимальная масса портфеля (ранца, сумки, рюкзака) с учебниками и учебными пособиями, тетрадями, школьно-письменными принадлежностями составляет 10% от массы тела обучающегося. Данный показатель может быть снижен с учетом уровня физического развития школьника, состояния его здоровья и удаленности школы от дома.
Как сообщается на сайте прокуратуры Московской области, соблюдение указанных требований к охране здоровья школьников возлагается на родителей, которые в соответствии с п. 1 ст. 63 Семейного кодекса Российской Федерации обязаны заботиться о здоровье своих детей.
Условия для детей с ограниченными возможностями
Все образовательные организации Московской области должны создавать условия для получения общего, профессионального и дополнительного образования детьми с ограниченными возможностями здоровья, в том числе в форме инклюзивного, дистанционного и надомного обучения, разработку и реализацию адаптированных и индивидуальных образовательных программ, содействие законным представителям в выборе форм и методов обучения, позволяющих наиболее полно реализовать интеллектуальный и творческий потенциал детей.
Инклюзивное образование в Подмосковье: доступная среда и подготовка кадров>>
Памятка «Оказание первой помощь при поражении электрическим током»
Как оказать первую помощь при поражении электрическим током
Поражение электротоком – это тот случай, когда человека обязательно нужно показать мед.работникам, даже если была грамотно оказана доврачебная помощь.
Ток может поразить внутренние органы, например, сердце или легкие, но сразу это заметно не будет, а проблемы проявятся позже. По этой же причине после сильного удара тока нужно постоянно наблюдать пострадавшего, проверять его самочувствие, при необходимости – немедленно показывать мед.персоналу.
Однако в наших силах принять меры по сохранению здоровья человека, по спасению его жизни после удара током, пока на место происшествия не прибыла скорая мед.помощь.
Алгоритм действий при оказании помощи пораженному электрическим током
Как можно быстрее вызовите мед. бригаду и приступайте к спасению человека. Лучше, если несколько человек будут заниматься этим одновременно. Алгоритм ваших действий:
бригаду и приступайте к спасению человека. Лучше, если несколько человек будут заниматься этим одновременно. Алгоритм ваших действий:
1. Если это возможно – сразу отключите электроустановку, до части которой дотронулся пострадавший. Нужно как можно скорее прекратить воздействие тока на него. От того, как долго ток будет действовать, будут зависеть и последствия. Самостоятельно разжать руку или отойти, когда бьет ток, человеку может быть очень сложно или невозможно, поэтому требуется срочная посторонняя помощь.
2. Когда отключить установку нет возможности, а человек держится за край кабеля или провода, кабель можно отрубить топором или другим подобным инструментом. У топора должна быть изолированная ручка – деревянная или пластиковая. Она обязательно должна быть сухой.
3. В электроустановках до 1000 Вольт допускается применение подручных средств (все они должны быть сухими и изолированными). Чтобы оттянуть человека, можно использовать деревянные палки, доски, сухие канаты.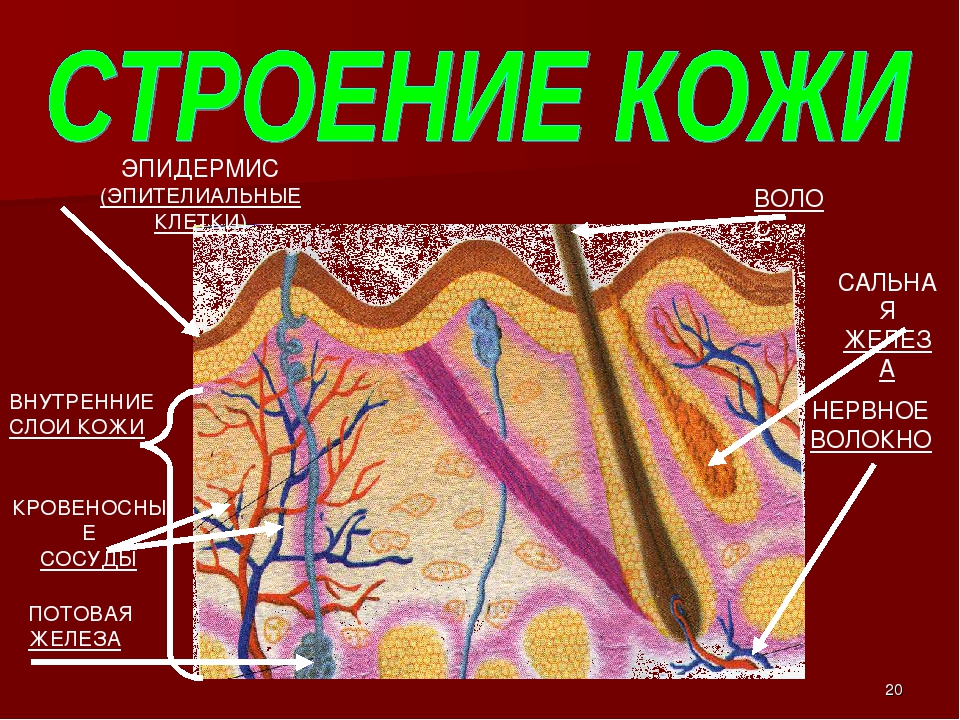 При условии, что у пораженного сухая одежда, можно потянуть за нее. При этом нужно быть внимательными и соблюдать меры предосторожности, заботиться о собственной безопасности: не прикасаться к самому человеку, его голой коже, а также к каким-либо предметам из металла и мокрым вещам.
При условии, что у пораженного сухая одежда, можно потянуть за нее. При этом нужно быть внимательными и соблюдать меры предосторожности, заботиться о собственной безопасности: не прикасаться к самому человеку, его голой коже, а также к каким-либо предметам из металла и мокрым вещам.
4. В электроустановках выше 1000 Вольт уже должны использоваться специальные инструменты и средства защиты: диэлектрические перчатки, ботинки или галоши, а также изолирующие штанги и щипцы.
Средства защиты от воздействия электрического тока
1. Под упавшего пораженного следует подложить сухую деревянную доску или фанеру.
2. Проверить наличие пульса и на запястье, и на шее.
3. Проверить зрачки: слишком широкие зрачки будут указывать на то, что кровоснабжение мозга пострадавшего сильно ухудшилось.
Далее действия зависят от того, в каком состоянии оказался человек после воздействия тока.
Оказание первой помощи при поражении электрическим током
Самые простые меры принимаются, если он в сознании.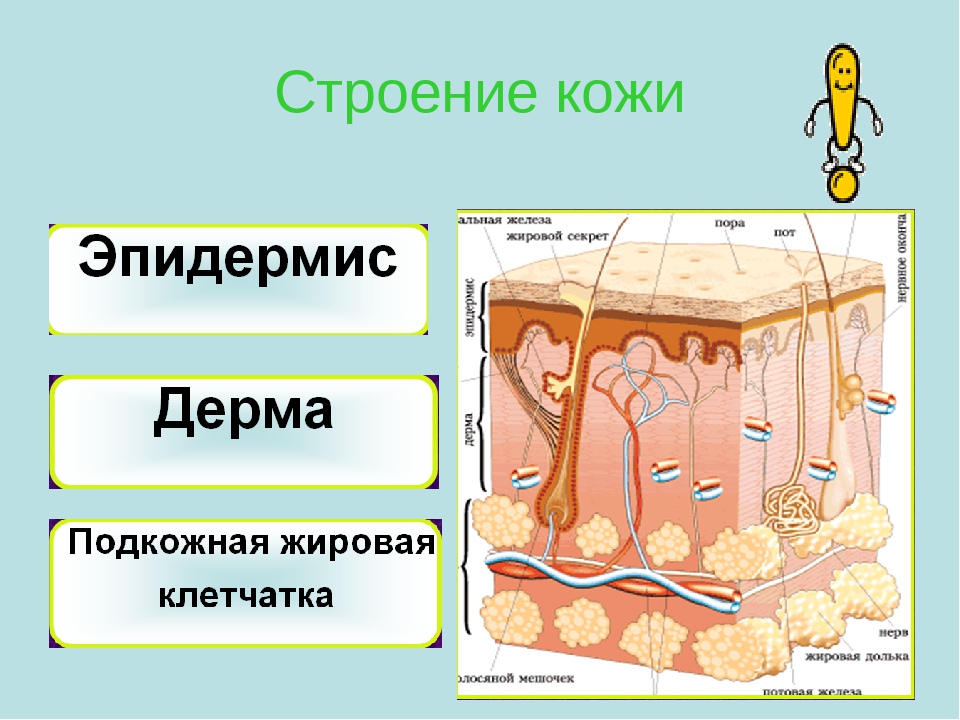 Пораженному нужно обеспечить покой. Пока вы дожидаетесь мед.помощи, уложите его как можно удобнее, укройте одеялом, постоянно проверяйте дыхание и пульс. При наличии ожогов, ушибов или переломов требуется оказание соответствующей доврачебной помощи. Если ничего подобного не обнаружено, не пытайтесь давать пострадавшему какие-то мед.препараты.
Пораженному нужно обеспечить покой. Пока вы дожидаетесь мед.помощи, уложите его как можно удобнее, укройте одеялом, постоянно проверяйте дыхание и пульс. При наличии ожогов, ушибов или переломов требуется оказание соответствующей доврачебной помощи. Если ничего подобного не обнаружено, не пытайтесь давать пострадавшему какие-то мед.препараты.
Человеку, потерявшему сознание, также нужен покой. Важно проверить, дышит ли он при этом. Необходимо уложить его на мягкую подстилку, расстегнуть на нем одежду, чтобы она не мешала дыханию, обеспечить доступ кислорода. Также меры спасения включают себя очищение рта: в его полости может скопиться кровь и слизь. До приезда мед.бригады нужно постараться согреть пострадавшего, а также следить за состоянием его дыхания.
Алгоритм действий включает в себя искусственное дыхание и непрямой массаж сердца, если пострадавший не подает признаков жизни или дышит прерывисто. Перед тем, как начать эти процедуры, как и в предыдущем случае, нужно освободить пораженного от стесняющей одежды, а также очистить его ротовую полость. Продолжать делать искусственное дыхание и массаж нужно до тех пор, пока человек не придет в себя или не приедет мед.персонал.
Описание Внешности на Английском
Если вы потеряли друга на людной вечеринке или ребенка в торговом центре, умение понятно описывать внешний вид очень пригодится. С помощью слов и синтаксических конструкций из этой статьи вы сможете самостоятельно составить рассказ о том, как выглядит человек.
Внешний вид по-английски — physical appearance или, менее формально, — looks. Если вы хотите узнать, как кто-то выглядит, используйте вопросы:
- What do you look like? — Как вы выглядите?
- How would you describe your physical appearance? — Как бы вы описали свою внешность?
- What does she/he look like? — Как она/он выглядит?
Не путайте вопросы о внешнем виде с вопросом «How would you describe yourself?» («Как бы вы себя охарактеризовали?»). Если вам задали такой вопрос, от вас ожидают описания личностных качеств, а не цвета глаз и волос.
Чтобы составить рассказ о внешности, можно придерживаться такого плана:
- цвет волос
- цвет глаз
- цвет кожи
- рост
- телосложение
- одежда (опционально)
Рассказ о внешности человека можно построить на двух основных речевых конструкциях.
Первая (классическая):
| Местоимение | + to be | + признак | ||||
| (глагол в личной форме) | (прилагательное) | |||||
| I | (Я) | am | tall. | |||
| He, she | (Он, она) | is | tall. | |||
| You, we, they | (Ты, вы, мы, они) | are | tall. | |||
Например:
| I am slender. | Я стройная. |
| She is short. | Она невысокая. |
| He is slightly overweight. | Он немного полноват. |
Вторая:
| Местоимение | + to have got | + признак | |
| (глагол в личной форме) | |||
| I | (Я) | have got (сокр.: ’ve got) | big blue eyes. |
| He, she | (Он, она) | has got (сокр.: ’s got) | big blue eyes. |
| You, we, they | (Ты, вы, мы, они) | have got (сокр.: ’ve got) | big blue eyes. |
Например:
| I have got red hair (или I’ve got red hair). | У меня рыжие волосы. |
| She has got green eyes (или She’s got green eyes). | У нее зеленые глаза. |
| He has got a beer belly (или He’s got a beer belly). | У него пивной животик. |
В статье «Названия частей человеческого тела на английском» вы найдете не только лексику по теме, но и полезные идиомы, связанные с телом.
Вместе с эпитетами, обозначающими внешний вид человека, можно использовать degree modifiers (обстоятельства степени):
- rather, quite, enough, pretty — довольно, достаточно
- very — очень
- too — слишком
| She is rather tall. | Она довольно высока. |
| He is very strong. | Он очень силен. |
Характеристика внешности — слова, описывающие человеческое тело
В английском языке существует огромное количество эпитетов, с помощью которых можно описать внешность человека.
Далее мы приводим довольно обширный перечень всевозможных характеристик человеческого тела. Используйте эту лексику для расширения словарного запаса: она пригодится вам не только в устной речи при необходимости описать себя или другого человека, но и при написании английского сочинения на тему «О себе», «О друге» или «Внешность человека».
Arms — Руки (от плеча до запястья)
| Bony | Костлявые |
| Lank, lanky | Худощавые |
| Long | Длинные |
| Rounded | Округлые |
| Short | Короткие |
| Well-shaped | Рельефные, хорошо очерченные |
Bearing — Осанка
| Bear oneself well | Иметь хорошую осанку |
| Posture | Осанка, поза |
| Stooping | Сутулый, согбенный |
| Upright | Прямая, правильная осанка |
| To hold one’s body upright | Иметь правильную осанку |
Build — Телосложение
| Anorexic | Анорексичный | Skinny, thin | Тощий, худой |
| Bonny | 1) Здоровый, цветущий 2) Милый | Slender | Стройный |
| Chubby | Пухлый, полный | Slight | Изящный, хрупкий |
| Delicate | Хрупкого телосложения | Slim, spare | Худощавый |
| Fat | Жирный, толстый | Small | Мелкого телосложения, тщедушный, субтильный |
| Graceful | Грациозный | Solid | Плотный |
| Heavy | Грузный | Stocky | Коренастый |
| Large | Массивный | Strong | Сильный |
| Neat | Стройный, изящный | Sturdy | Крепкий, дюжий |
| Overweight | С избыточным весом | Tubby | Пузатый, толстый |
| Plump | Полный, пухлый | Underweight | С недостаточным весом |
| Powerful | Мощный | Well-built, muscular | Хорошо сложенный, мускулистый |
Полного человека лучше не называть fat или tubby: это может его обидеть; лучше сказать, что он имеет лишний вес (a bit overweight).
Cheekbones — Скулы
| Perfect | Идеальные |
| Prominent | Выступающие |
| Sculptured | Скульптурные |
| with high cheekbones | Скуластый, широкоскулый |
Cheeks — Щеки
| Ashen | Мертвенно бледные |
| Blushing | Пылающие, залитые румянцем |
| Chubby | Полные |
| Flushed | Раскрасневшиеся |
| Hamster, hamster-like | Как у хомяка |
| Hollow, sunken | Впалые, ввалившиеся |
| Pale, pallid | Бледные |
| Pasty | Мертвенно бледные |
| Pink, rosy | Розовые |
| Plump, stubby | Пухлые |
| Rouged | Нарумяненные |
| Ruddy | Румяные, красные, свежие |
| Scarlet | Алые |
| Smooth | Гладкие |
| Soft | Мягкие, нежные |
| Tear-stained | Заплаканные |
| Unshaven | Небритые |
| Wet | Мокрые |
| Wide | Широкие |
| with creases, with wrinkles | Морщинистые |
| with dimples | С ямочками |
Chin — Подбородок
| Clean-shaven | Гладко выбритый |
| Decided | Решительный |
| Double | Раздвоенный |
| Massive | Массивный |
| Pointed | Заостренный |
| Protruding | Выступающий |
| Round | Округлый |
Constitution — Конституция, комплекция
| Delicate | Нежный |
| Feeble, weak | Слабый, хилый |
| Frail | Хрупкий |
| Iron | Железный |
| Rugged, vigorous | Выносливый |
| Strong | Сильный |
Ears — Уши
| Earlobe | Мочка уха |
| Large | Большие |
| Lop-eared | Лопоухий |
| Pointed, pointy | Заостренные |
| Small | Маленькие |
Eyelashes — Ресницы
| Artificial, false, «falsies» | Искусственные (накладные) |
| Curling | Подкрученные, загнутые на концах |
| Long | Длинные |
| Short | Короткие |
| Straight | Прямые |
| Thick | Густые |
| Thin | Жидкие |
Eyebrows — Брови
| Amused | Удивленно приподнятые |
| Arched | Дугообразные |
| Bushy, thick | Густые |
| Dark | Темные |
| Derisive, mocking, quizzical | Насмешливые |
| Enquiring, querying, questioning | Вопросительно приподнятые |
| Jet-black | Черные как смоль |
| Lifted, raised | Приподнятые |
| Pencilled | Подведенные карандашом |
| Plucked | Выщипанные |
| Raise (knit) one’s brows | Поднять брови |
| Sardonic | Сардонически приподнятые |
| Skeptical | Скептически приподнятые |
| Shaggy | Лохматые |
| Shaped | Оформленные (выщипанные) |
| Straight | Прямые |
| Surprised | Удивленные |
| Thick | Густые |
| Thin | Жидкие, тонкие |
| Unkempt | Неухоженные |
| Well-marked | Выразительные |
Eyelids — Веки
| Big, large | Большие |
| Closed | Смеженные, сомкнутые |
| Drooping | Поникшие |
| Half-closed | Полузакрытые |
| Heavy | Тяжелые |
| Hooded, lowered | Нависающие |
| Lower | Нижнее (веко) |
| Swollen with tears | Опухшие от слез |
| Upper | Верхнее (веко) |
Eyes — Глаза
| Almond-shaped | Миндалевидные | Keen | Проницательные |
| Angry | Сердитые | Limpid | Прозрачные |
| Anxious | Беспокойные | Liquid | Водянистые |
| Baggy | С мешками под глазами | Lively | Живые |
| Beady | Глаза-бусинки | Luminous | Светящиеся |
| Big | Большие | Lustrous | Блестящие |
| Bleary | Мутные | Mad | Безумные |
| Bloodshot | Налитые кровью | Mesmerizing | Гипнотизирующие, завораживающие |
| Blue | Синие, голубые | Misty | Затуманенные |
| Bright | Яркие | Moist | Влажные |
| Brilliant | Блестящие | Narrow | Узкие |
| Brown | Коричневые | Narrowed | Сузившиеся |
| Bulging | Выпуклые | Pale | Бесцветные |
| Clear | Ясные | Penetrating, piercing | Пронзительные |
| Close-set | Близко посаженные | Piggy | Поросячьи |
| Cloudy | Затуманенные | Protuberant | Выпуклые |
| Cold | Холодные | Prying | Любопытные |
| Compassionate | Сострадательные | Puffy | Опухшие |
| Crinkly | В морщинках | Red-rimmed | Покрасневшие, воспаленные |
| Cross-eyed | Косоглазый | Rheumy | Слезящиеся |
| Cruel | Жестокий (взгляд) | Round | Круглые |
| Curious | Любопытные | Sad | Грустные, печальные |
| Dark | Темные | Sharp | Острый (глаз), хорошее (зрение) |
| Dark-ringed | С темными кругами (под глазами) | Short-sighted | Близорукие |
| Dazed | Ошеломленные | Shrewd | Проницательные |
| Deep | Глубокие | Sightless | Незрячие |
| Deep-set, downcast | Глубоко посаженные | Slanted | Раскосые |
| Dry | Сухие | Sleepy | Сонные |
| Enormous | Огромные | Small | Маленькие |
| Exhausted | Измученные | Soft | Нежные |
| Expressionless, hollow, vacant | Пустые, ничего не выражающие | Solemn | Торжественные |
| Expressive | Выразительные, многозначительные, красноречивые | Sorrowful | Грустные, печальные |
| Fierce | Пронзительные | Soulful | Проникновенные |
| Firm | Твердый (взгляд) | Sparkling | Оживленные, искрящиеся |
| Full | Налитые кровью | Squinty | Узкоглазый (пренебр.) |
| Gentle | Нежные | Staring | Широко раскрытые (глаза), пристальный (взгляд) |
| Glassy, glazed | Безжизненные, стеклянные, остекленевший (взгляд) | Starry | Лучистые |
| Glasz | Разноцветные (сине-серо-зелено- коричневые) | Steely | Стальной (взгляд) |
| Golden | Золотистые | Sunken | Ввалившиеся |
| Gray | Серые | Swollen | Опухшие |
| Greedy | Жадные | Sympathetic | Сочувствующий (взгляд) |
| Green | Зеленые | Tear-filled, tearful | Полные слез |
| Half-closed | Полузакрытые | Tired, weary | Уставшие |
| Hazel | Ореховые | Twinkling | Мерцающие |
| Heavy-lidded | С тяжелыми веками | Unblinking | Немигающие |
| Hooded | С нависающими веками | Unfocused | Бесцельный (взгляд) |
| Huge | Огромные | Unseeing | Невидящие |
| Hungry | Голодные | Velvety | Бархатный (взгляд) |
| Intelligent | Умные | Warm | Теплый (взгляд) |
| Keen | Проницательные | Watery | Водянистые |
| Large | Большие | Wide-apart, wide-set | Широко расставленные |
| Laughing | Смеющиеся | Wild | Дикие, безумные |
-eyed в составе слов соответствует -глазый, -окий: black- eyed — черноглазый, green-eyed — зеленоокая и т.п.:
Beware of jealousy, my lord! Shakespeare, “Othello”, Act 3, Scene 3 | Берегитесь ревности, синьор. Шекспир, «Отелло», акт 3, сцена 3 |
- Black eye означает синяк под глазом.
Face (shape) — Лицо (форма)
| angular | угловатое | round | круглое |
| heart-shaped | «сердечком» | square | квадратное |
| long | удлиненное | thin | худое, исхудавшее |
| oval | овальное |
Face (complexion) — Лицо (цвет лица, кожа)
| baby-soft | нежная, как у ребенка | rough | грубая |
| callused | загрубевшая | ruddy | (о лице) румяное |
| dark | темная | sallow | землистого цвета |
| dry | сухая | silky | шелковистая |
| fair | светлая | smooth | гладкая |
| florid | цветущая | soft | нежная |
| freckled | веснушчатая | spotless | здоровая |
| glowing | сияющая | sunburnt | загорелая, обожженная солнцем |
| olive | оливкового цвета | swarthy | смуглая |
| pale | бледная | tanned | загорелая |
| paper-thin | тонкая, как бумага | translucent | прозрачная |
| pasty | бледная как смерть | wrinkled | морщинистая |
| rosy | розовая | ||
| -faced в составе слова соответствует -лицый: broad-faced (широколицый) и т.д. | |||
Figure — Фигура
| bony | костлявая | plump | пухлая |
| clumsy | неуклюжая | shapeless | бесформенная |
| flawed | небезупречная | slender | стройная |
| graceful | изящная | slight | хрупкая, изящная |
| ill-made | некрасивая, непропорциональная | slim | стройная |
| lean | поджарая, сухопарая | stocky | коренастая |
| lithe | гибкая | stout | полная, плотная |
| neat | аккуратная | superb | превосходная |
| ordinary | обыкновенная, ничем не примечательная | tattooed | (о теле) татуированное |
| paunchy | пузатая, «с брюшком» | well-fed | откормленная |
| perfect, great | прекрасная, великолепная | well-made | ладная |
Fingers — Пальцы
| arthritic | артритные | lean | худые |
| blunt | тупые | little finger | мизинец |
| bony | костлявые | middle finger | средний палец |
| capable | умелые | nerveless | слабые |
| chubby | пухлые | nervous | нервные |
| clumsy | неуклюжие | nimble | ловкие |
| deft | ловкие | plump | пухлые |
| delicate | нежные, изящные | podgy | короткие и толстые |
| dirty, filthy | грязные | ring finger | безымянный палец |
| elegant | изящные | skillful, skilled | искусные, умелые |
| fat | толстые | skinny | худые |
| finger cushion, finger-pad | подушечки пальцев | slim | тонкий |
| finger nail | ноготь | spatulate | лопатообразные |
| forefinger | указательный палец | square-tipped | с квадратными кончиками |
| gentle | нежные | sticky | липкие |
| gnarled | корявые | toe | палец ноги |
| grubby | неопрятные | thumb | большой палец |
| index finger | указательный палец |
Forehead — Лоб
| broad | широкий | wide | широкий |
| domed | куполообразный | narrow | узкий |
| furrowed | изрытый, изборожденный морщинами | retreating, sloping | покатый |
| high | высокий | wrinkled | морщинистый |
| low | низкий | large | большой |
| smooth | гладкий | small | маленький |
Gait — Походка
| awkward | неуклюжая | steady | уверенная |
| brisk | бодрая | stride with a firm step | шагать твердым шагом |
| careful, delicate | осторожная | strolling | прогуливающаяся |
| firm | твердая | stumbling | спотыкающаяся |
| gingerly | робкая, опасливая | swaggering | важная |
| halting | неровная | swaying | шатающаяся |
| heavy | тяжелая | unsteady | неверная |
| light | легкая | waddling | вразвалочку |
| measured footstep | размеренный шаг | walk with a drag | волочить ногу |
| quick, rapid | быстрая | walk with a droop | горбиться при ходьбе, ходить сутулясь |
| shuffling | шаркающая | walk with a limp | хромать |
| slow | медленная | walk with a staff (stick) | идти, опираясь на палку (трость) |
| smart | энергичная |
General appearance — Внешний вид (в общем) | |||
| appearance, looks | внешность | nice | милый |
| attractive | привлекательный | not look oneself | быть непохожим на себя |
| beautiful | красивый | plain | простоватый |
| charming | очаровательный | pleasant-looking | приятный |
| common | обычный | posh, gorgeous | роскошная, шикарная (о женщине) |
| eyesore | уродливый | pretty | симпатичная, хорошенькая (о девушке, женщине) |
| family likeness | фамильное сходство | repulsive | отталкивающий |
| good-looking | симпатичный | resemble | напоминать |
| handsome | красивый (о мужчине) | take after somebody | быть похожим на (кого-л.) |
| impressive | впечатляющий | ugly | уродливый |
| look | внешний вид, образ | unattractive | непривлекательный |
| look like | выглядеть как (кто-л.), быть похожим на (кого-л.) | unimpressive | невыразительный |
| look one’s age | выглядеть на свой возраст | unpleasant-looking | неприятный |
| look one’s best | выглядеть наилучшим образом | unsightly | неприглядный |
| look wretched | выглядеть несчастным | usual | обычный |
| lovely | прекрасный, очаровательный | ||
Hair — Волосы
| abundant | густые, роскошные | long | длинные |
| baby-fine | тонкие, как у ребенка | lustrous | блестящие |
| bald | лысый | luxuriant | пышные |
| bald spot | лысина | mop of hair | копна волос |
| balding | лысеющий | neatly-combed | аккуратно зачесанные |
| bushy | густые | parted | разделенные на пробор |
| coarse | жесткие | permed | после химической завивки |
| crisp | кудрявые, курчавые | scraggly | всклокоченные |
| cropped | подстриженные | shaved | обритые |
| curly | вьющиеся | short | короткие |
| dishevelled | растрепанные | silky | шелковистые |
| dyed | окрашенные | spiky | остроконечные (пряди) |
| fair | светлые | stiff | жесткие |
| fine | тонкие | straight | прямые |
| fine crop of hair, magnificent bush/head of hair | роскошная шевелюра | thick | густые |
| thin | тонкие, жидкие | ||
| flat | приглаженные | wavy | волнистые |
| flaxen | льняные | weaved | заплетенные |
| full | густые | wiry (stiff) | похожие на проволоку ( жесткие) |
| head of curly hair | кудрявая шевелюра | wispy | тонкие |
| limp | редкие |
Hair colors — Цвет волос
| ash-blond | пепельный блондин | gray, grey(-ing) | седой, седеющий |
| auburn | красно-коричневый | highlighted | с цветовыми бликами, с контрастными прядями |
| black | черный | honey-blond | медового цвета |
| bleached | обесцвеченный | jet-black | черные как смоль |
| blond(e) | блондин(ка), белокурый цвет волос | red | рыжие |
| brown | коричневый | reddish | рыжеватые |
| brunet(te) | брюнет(ка) | redhead | рыжеволосый(-ая) |
| chestnut, chestnut-brown | каштановый | salt-and-pepper | черные с проседью, «соль с перцем» |
| dark-haired | темноволосый | silvery | серебристые |
| golden | золотистые | strawberry-blond | пшеничного цвета, блондин с рыжеватым оттенком |
| golden-blond | золотистый блондин | white | белые |
Hair (men’s hairstyle & haircut) — Волосы (мужские стрижки)
| bald patch | плешь | bald-head | лысый |
| closely-cropped | коротко подстриженные (волосы) | crew-cut, buzz cut | стрижка «ежиком» |
| dreadlocks | прическа «африканские косички» | ||
| flat top | стрижка «площадка» | partly bald-head | с залысинами, с проплешинами |
Hair (women’s hairstyle) — Волосы (женские прически)
| bangs | челка |
| bobbed hair | стрижка «боб» |
| braids, pigtails, plaits | косы, косички |
| brush one’s hair | причесываться |
| bun, knot of hair, tuft of hair | пучок волос |
| bunches | «хвостики», afro-bunches — африканские косички |
| comb one’s hair | расчесывать волосы |
| curled hair | завитые волосы |
| hair-cut | стрижка |
| hairdo, hairstyle | прическа |
| lock, ringlet | локон |
| pinned-up hair, swept-up hair | заколотые, подобранные вверх волосы |
| ponytail | прическа «конский хвост» |
| shoulder-long | волосы длиной до плеч |
| slicked down | «прилизанные» волосы |
| strand of hair, wisp of hair | прядь волос |
| swept-back hair | волосы, зачесанные назад |
| trimmed hair | подстриженные волосы |
| twist | прическа «ракушка» или «завиток» |
| She has long hair. | She has short hair. | She has short hair. | She has medium length hair. | She has medium length hair. | He has no hair. |
| She has long, dark hair. | She has short, black hair. | She has short, fair hair. | She has medium length, blonde hair. | She has medium length, red hair. | |
| She has long, straight, dark hair. | She has short, straight, black hair. | She has short, straight, fair hair. | She has medium length, curly, blonde hair. | She has medium length, wavy, red hair. | |
| Her hair is long, straight and dark. | Her hair is short, straight and black. | Her hair is short, straight and fair. | Her hair is medium length, curly and blonde. | Her hair is medium length, wavy and red. | He is bald. |
Hair (facial) — Усы, бороды, бакенбарды
| beard | борода | military moustache | военные усики |
| bearded | бородатый | neat moustache | аккуратные усики |
| bushy | заросший | painter’s brush moustache | усы «кисть художника» |
| clean shaven | чисто выбритый | pencil moustache | усы-«карандаш» |
| clipped | подстриженные (усы, борода) | pyramidal moustache | «пирамидальные» усы |
| curly | вьющиеся | side whiskers, sideboards, sideburns | бакенбарды, бачки |
| drooping, droopy | поникшие (усы) | smooth cheeks | гладкие щеки |
| five o’clock shadow (stubble) | щетина | stubble beard | щетина |
| full beard | длинная борода | stubby | коренастый |
| fu manchu moustache | «фу манчи», «маньчжурские» усы | thick | густая (борода и т.д.) |
| goatee | бородка клинышком, эспаньолка (букв. «козлик») | thin | жидкая (борода и т.д.) |
| toothbrush | усики в стиле «зубная щетка» | ||
| handlebar moustache | подкрученные вверх длинные усы, «велосипедный руль» | unshaven | небритый |
| walrus moustache | длинные свисающие «моржовые» усы | ||
| horseshoe moustache | усы подковой | waxed moustache | напомаженные, набриолиненные усы |
| imperial moustache | имперские усы | with a pointed beard and moustache | с остроконечной бородкой и усами |
| lampshade moustache | усы «абажур» | ||
| Примечание: в отличие от русского языка, слово moustache («усы») в английском языке имеет единственное число: He has an elegant moustache. — У него элегантные усики. | |||
Hands — Руки (ладони, пальцы)
| arthritic | артритные | nervous | нервные |
| beautiful | красивые | open | раскрытая (ладонь) |
| capable, expert, practiced, skilled, skillful | умелые, искусные | outspread | распростертые |
| careless | небрежные | plump | пухлые |
| clammy | липкие | ringed | унизанные кольцами и перстнями |
| clean | чистые | ringless | без колец |
| clumsy | неуклюжие | shaking, trembling | дрожащие |
| coarse, rough | грубые | short | короткие |
| delicate | нежные, изящные | soft | мягкие |
| dirty | грязные | steady | уверенные |
| fat | жирные | sticky | липкие |
| filthy | грязные | sweaty | потные |
| firm | твердые | thick | толстые |
| frail | хрупкие | unsteady | неуверенные |
| gnarled | корявые | unwashed | немытые |
| greasy | засаленные | well-manicured | ухоженные |
| grubby | неряшливые | work-reddened | покрасневшие от работы |
| hairy | волосатые | work-roughened | огрубевшие от работы |
| limp | вялые | work-worn | натруженные |
Head & neck — Голова и шея
| big | большая (голова) |
| hold one’s head high | с высоко поднятой головой |
| round | круглая (голова) |
| slender | изящная, грациозная (шея) |
| small | маленькая (голова) |
| square | квадратная (голова) |
| thick | толстая (шея) |
| thin | тонкая (шея) |
Jaws — Челюсти
| artificial | искусственная | jutting, lantern, thrusting | выступающая |
| bottom | нижняя | lower | нижняя |
| broken | сломанная | pointed | заостренная |
| clenched | сомкнутые (челюсти) | (to) set jaw | стиснуть зубы |
| dislocated | вывихнутая | slack-jawed | с отвисшей челюстью |
| false | фальшивая | square | квадратная |
| firm | твердая | strong | сильная |
| fractured | сломанная | top, upper | верхняя |
Legs — Ноги
| bandy (legs) | О-образные | muscled, muscular, shapely | мускулистые |
| beautiful | красивые | ||
| bow-shaped | дугообразные | narrow | узкие (о ступне) |
| crooked | изогнутые | powerful | мощные |
| good | хорошие | fine, slender | стройные |
| hairy | волосатые | short | короткие |
| lank | худощавые | narrow | узкие (о ступне) |
| large, stout, stumpy | массивные | small | маленькие (ступни) |
| limb | конечность | strong | сильные |
| long | длинные | thin | тонкие |
Lips — Губы
| bottom, lower | нижняя | painted | накрашенные |
| chapped, cracked | потрескавшиеся | sensual, sensuous | чувственные |
| compressed | сжатые | thick | толстые |
| dry | сухие | thin | тонкие |
| firm | твердые | top, upper | верхняя |
| fleshy | мясистые | weak-willed | безвольные |
| full | полные | well-cut, well-shaped | хорошо очерченные |
| moist | влажные |
Marks — Родинки, морщины, шрамы и др.
| crow’s feet | «гусиные лапки» (мелкие лучеобразные морщинки) |
| dimple (with a dimple in the chin) | ямочка (с ямочкой на подбородке) |
| mole (with a mole) | родинка (с родинкой) |
| scar | шрам |
| spot (pimple) | прыщ |
| wrinkle | морщина |
Mouth — Рот
| beautiful | красивый | lipless | безгубый |
| big | большой | lopsided | асимметричный |
| cavernous | похожий на пещеру | lovely | очаровательный |
| chiseled | рельефный | pretty | хорошенький |
| cruel | жестокий | sensual, sensuous | чувственный |
| dry | сухой | small | маленький |
| enormous, huge | огромный | soft | нежный |
| firm, hard | твердо сжатый | stern | сурово сжатый |
| full | полный | strong, strong-willed | волевой |
| full-lipped | с полными губами | thin | тонкий |
| gaping | зияющий | tight | плотно сжатый |
| half-open | полуоткрытый | toothless | беззубый |
| handsome | красивый | well-shaped | хорошо очерченный |
| hungry | голодный | wet | влажный |
| large | большой | wide | широкий |
Nose — Нос
| aquiline | орлиный | pink | розовый |
| aristocratic | аристократический | pointed | заостренный |
| beaky | клювообразный | prominent | выступающий, выдающийся |
| big, large | большой | red | красный |
| bridge of the nose | переносица | Roman | римский |
| bulbous | нос луковкой | sharp | острый |
| crooked, curved | изогнутый | shiny | блестящий |
| elegant | элегантный, изящный | small | маленький |
| enormous, huge | огромный | snub | курносый |
| flat | плоский | straight | прямой |
| flattened | сплюснутый | stubby | короткий и толстый |
| fleshy | мясистый | thin | тонкий |
| hooked | нос крючком | tiny | крошечный |
| little | маленький | tip of the nose | кончик носа |
| long | долго | tip-tilted | со вздернутым кончиком |
| narrow | узкий | ||
| nostrils | ноздри | turned up, upturned | вздернутый |
Shoulders — Плечи
| bare | обнаженные | muscled, muscular | мускулистые |
| big | большие | narrow | узкие |
| bony | костлявые | powerful | мощные |
| bowed | сутулые | round | округлые |
| broad | широкие | shapely | красивые, правильной формы |
| delicate | хрупкие | slim, thin | худые |
| great | великолепные | sloping | покатые |
| huge | огромные | square | квадратные |
| manly | мужественные | strong | сильные |
| massive | массивные | wide | широкие |
Stature, height & shape — Телосложение, рост
| be 6 feet high | быть 6 футов ростом |
| be of average / normal / medium height | быть среднего / нормального роста |
| diminutive | миниатюрный |
| imposing | импозантный |
| short | маленького роста |
| tall | высокий |
| waist (be slim at the waist) | талия (иметь тонкую талию) |
Teeth — Зубы
| baby | детские | needle-sharp | острые, как иглы |
| back | задние | pearl-like, pearl-white | жемчужно-белые |
| big, large | большие | ||
| bottom | нижние | perfect | идеальные |
| broken | сломанные | prominent, protruding | выступающие |
| crooked, twisted | кривые | razor-sharp | острые как бритва |
| decayed | плохие | rotten | гнилые |
| enormous, huge | огромные | sharp | острые |
| even | ровные | small | небольшие |
| false, artificial | вставные, искусственные | sparse | редкие |
| front | передние | splendid | великолепные |
| gappy | редкие, с промежутками | sticking out | торчащие |
| gold | золотые | straight | прямые |
| good | хорошие | strong | крепкие |
| healthy | здоровые | top | верхние |
| imperfect | неидеальные | uneven | неровные |
| jagged | зазубренные | white | белые |
| milk | молочные | wisdom tooth | зуб мудрости |
| misshapen | деформированные | wobbly, loose tooth | шатающийся зуб |
| missing | отсутствующие | yellow | желтые |
Указывать рост в футах и дюймах, а вес — в фунтах придется, если ваш собеседник из Англии и США. В этих странах используется традиционная английская система мер.
Примеры описания внешностиЧаще всего приходится описывать чужую внешность, когда мы кого-то ищем или рассказываем о человеке, которого собеседник не знает. Мы подготовили несколько примеров из жизни, по образцу которых вы можете составить собственные рассказы о внешнем виде знакомого человека.
Поиск человека в соцсетиI’m looking for a girl I saw yesterday on the train. Your seat was across mine and you were reading all the time. You’re short and slender, with shoulder-length black hair and charming green eyes, and you’ve got left brow piercing. You were dressed in white tank top and blue skinny jeans. If you’re reading this, please hit me up. | Ищу девушку, которую видел вчера в поезде. Ты сидела напротив меня и читала книгу. Ты невысокая и стройная, с черными волосами до плеч и очаровательными зелеными глазами, и у тебя на левой брови пирсинг. Была одета в белую майку и синие узкие джинсы. Если ты читаешь это, свяжись со мной, пожалуйста. |
A five-years old boy called Andrei is lost at Aura Mall. He is dressed in a green coat and striped beanie. The boy might look short for his age. He’s got brown eyes, chestnut hair and a large mole on his forehead. Andrei’s parents are waiting for him at the information desk. | В торговом центре «Аура» потерялся мальчик пяти лет по имени Андрей. Одет в зеленую куртку и полосатую шапочку. Мальчик выглядит невысоким для своего возраста. У него карие глаза, каштановые волосы и большая родинка на лбу. Родители ждут Андрея возле стойки информации. |
Mike: So, what does Sarah’s new boyfriend look like? Angel: Well, I’d say he’s got a very manly appearance. He has a bushy beard, a perfect posture and a firm glance. Mike: What about his body, is he in good shape? Angel: Oh yes, he is! It seems like he’s naturally well-built and broad-shouldered, and he’s obviously working out a lot. He’s so ripped! Mike: I see. Sounds like Sarah is doing well. | Майк: Ну и как выглядит новый парень Сары? Энджел: Ну, я бы сказала, что у него весьма мужественная внешность. Густая борода, идеальная осанка и твердый взгляд. Майк: Что насчет фигуры, он в хорошей форме? Энджел: Еще как! Похоже, что у него от природы хорошее телосложение и широкие плечи, и он явно ходит в спортзал. Он такой накачанный! Майк: Понятно. Похоже, у Сары все хорошо. |
Sam: I never knew you have a sister! Are you twins? Keira: No, she’s two years older than me. And she actually looks nothing like me. Sam: Really? What does she look like? Keira: Well, she’s a blonde like our dad, and I have our mom’s reddish hair. She’s also a bit taller than me and quite slim, while I’m definitely more chubby. Sam: Do you think I would know she’s your sister if I saw her? Keira: I don’t think so. Even our facial features are different. She has pale spotless skin, while mine is freckled. Her lips are thicker than mine and her nose is wider. But we both have our mom’s blue eyes. | Сэм: Я не знал, что у тебя есть сестра! Вы двойняшки? Кира: Нет, она на два года старше меня. И на самом деле она совсем на меня непохожа. Сэм: Правда? Как она выглядит? Кира: Ну, у нее светлые волосы, как у нашего отца, а у меня они рыжеватые, как у мамы. Она немного выше меня и довольно стройная, а я определенно более полненькая. Сэм: Как ты думаешь, я бы понял, что это твоя сестра, если бы ее увидел? Кира: Вряд ли. У нас даже черты лица разные. У нее кожа бледная и безупречная, а у меня веснушки. У нее более пухлые губы и более широкий нос, чем у меня. Но у нас обеих мамины голубые глаза. |
- Базовый рассказ о внешности состоит из описания цвета глаз, волос и кожи, а также роста и телосложения.
- Полезные речевые конструкции: I am… / she is… и I’ve got… / he’s got… Например: «I am slender» («Я высокая»), «She’s got green eyes» («У нее зеленые глаза»).
- При описании внешности пригодятся обстоятельства степени: rather, quite, enough, pretty — довольно, достаточно, very — очень и too — слишком. Суффикс -ish тоже поможет выразить степень, например: reddish — рыжеватый.
На занятиях в Skyeng вы потренируетесь описывать людей по фотографиям и использовать разговорные конструкции для рассказа о внешности. Оставьте заявку, чтобы бесплатно посетить вводный урок в самой большой онлайн-школе английского языка в Европе.
Читаем дальше:
Как рассказать о возрасте человека: Времена… жизни
Как описать наряд человека:
Словарь шопоголика: глоссарий предметов одежды, фасонов и материалов на английском
Гид-путеводитель по миру обуви
Рак печени: лечение, симптомы, диагностика рака печени 4 степени
Сегодня основной метод лечения злокачественных новообразований печени при первичных опухолях и метастазах колоректального рака – анатомическая лапароскопическая резекция печени. Такая методика используется в странах Западной Европы, в Израиле, в США и в LISOD. При метастазах других видов рака применяют атипичную резекцию, радиочастотную абляцию.
Рак печени или печеночноклеточный рак – злокачественная опухоль, которая локализуется в печени. Новообразование происходит из клеток печени или является метастазом другой (первичной) опухоли. Метастазы в печени развиваются намного чаще, чем первичные опухоли. Это связано с характером кровообращения и функцией печени в организме. Метастазы злокачественного новообразования – это тяжелое осложнение, которое более опасно, чем сама первичная опухоль. Метастатический рак выявляют как метастазы злокачественной опухоли при ее первичной локализации в других органах.
Первичный рак печени имеет следующие типы: гепатоцеллюлярная карцинома, ангиопластическая саркома (ангиосаркома), гепатобалстома и гемангиосаркома, холангиокарционома.
Саркома печени — редкое и опасное заболевание. Очень быстро происходит развитие опухоли, распространение на соседние органы. Особенностью сарком является их возникновение преимущественно у людей молодого возраста и у детей. В частности, ангиосаркома печени – это редкая форма злокачественной опухоли. Новообразование имеет агрессивное течение, проявляется инвазивным, очень быстрым ростом, метастазирует.
Холангиокрацинома или рак желчного протока может развиться как в области печени, так и за пределами органа. Опухоль может образоваться в любой части протока. В зависимости от локализации образования существуют три группы: рак внутрипеченочных протоков, рак желчного протока в области ворот печени, рак дистальных желчных протоков.
Причины и механизм развития заболевания еще недостаточно изучены. Исследовалось влияние географического положения, климата, рациона питания, некоторых медицинских препаратов. У больных алкоголизмом первичный печеночноклеточный рак нередко развивается на фоне цирроза. Рак печени может обнаружиться в любом возрасте, чаще им заболевают после 40 лет.
Диагностика
В LISOD для диагностики рака печени применяют современные методы, соответствующие международным медицинским стандартам.
Ультразвуковое исследование (УЗИ) позволяет обнаружить опухоль и в некоторых случаях определить ее тип.
Биопсия опухоли является наиболее достоверным методом диагностики рака печени. Как правило, применяют тонкую длинную иглу, которую вводят через кожу в печень в область опухоли под контролем УЗИ-аппарата. Если во время исследования участка новообразования под микроскопом обнаруживают раковые клетки, диагноз рака печени считается подтвержденным.
Компьютерная томография (КТ) очень эффективна при диагностике опухолей печени, позволяет обнаружить даже маленькие новообразования, незаметные на УЗИ. В LISOD для улучшения изображения проводится КТ с контрастированием – внутривенно вводится контрастное вещество, что дает возможность специалистам изучить расположение сосудов в печени. Во время компьютерной томографии аппарат получает изображение тонких срезов, что позволяет специалистам тщательно обследовать структуру органа и выявить даже небольшие опухоли.
Лапароскопия. Лапароскопический метод позволяет поставить точный и правильный диагноз. Метод щадящий, быстрый и безболезненный. Через небольшой разрез, под короткодействующим наркозом, врач вводит специальный прибор в брюшную полость, осматривает опухоль (на мониторе) и берет кусочек ткани на исследование.
Исследование крови. Определение уровней альфа-фетопротеина (АФП) в крови полезно как на этапе диагностики опухолей печени, так и после лечения для контроля эффективности терапии и возможного рецидива заболевания.
Лечение
Полную информацию о диагностике и лечении этого вида рака Вам предоставят консультанты Информационной службы LISOD:
- 0-800-500-110 (бесплатно для звонков
со стационарных телефонов по Украине) - или +38 044 520 94 00 – ежедневно
с 08:00 до 20:00.
Прежде всего, врачи уточняют диагноз опухоли и степень ее распространения. План лечения разрабатывается на междисциплинарной онкологической конференции.
Если опухоль операбельна, то начинается подготовка к серьезному хирургическому вмешательству. Удаление метастазов в печени – эффективный метод лечения. Печень «умеет» регенерироваться, и удаленная часть постепенно восстанавливается. Таким образом, ведущим методом остается радикальное оперативное вмешательство (например, гемигепатэктомия или атипичные резекции печени) в сочетании с последующей химиотерапией.
Радиотерапия также применяется в лечении рака печени, лечении метастазов в печень, так как снижает темпы роста опухоли. Радиотерпия – эффективный метод лечения метастазов в печени. Радиотерапия может быть использована в сочетании с хирургическим лечением либо с химиотерапией. Эмболизация печеночной артерии подразумевает блокировку артериальной крови, которая переносится к карциноидным опухолям с последующим проведением химиотерапии для уменьшения размеров оставшихся очагов.
Ранее выявление болезни является залогом успешного лечения. Операция, проведенная на ранних стадиях рака печени, как правило, дает хорошие результаты.
Симптомы
Рак печени симптомы проявляет, как правило, на фоне хронических болезней (вирусный гепатит, цирроз и т.д.). В результате, на признаки уже существующего недуга накладываются новые.
Появление боли в животе или ее усиление – симптом рака, часто свидетельствующий о больших размерах новообразования или его распространения за пределы печени.
Ощущение тяжести в правом подреберье может быть признаком рака печени.
Повышенная температура тела (выше 37.5 градусов Цельсия), которая держится длительное время и не объясняется другими причинами.
Появление водянки (асцит) – это скопление в брюшной полости жидкости, возникающее при раке печени и циррозе. Т.е. появление асцита у больного с циррозом может быть как осложнением основной болезни, так и признаком рака печени.
Желтуха – пожелтение склер глаз, слизистых оболочек и кожи тела. Возникает при разных состояниях, но, в том числе, при циррозе и при раке печени. Желтуха у больного может проявляться как осложнение цирроза, но так же может быть признаком рака печени.
Вздутие живота, снижение массы тела, отсутствие аппетита, выраженная слабость – это симптомы рака печени, но могут встречаться и при других болезнях.
Факторы риска
Известны факторы риска, которые могут способствовать развитию рака печени.
Пол. Мужчины болеют чаще, чем женщины. Возможно, это связано с большим употреблением ими алкоголя.
Заболевания печени. Хроническая инфекция (гепатит С или В) – очень значимый фактор риска. Есть некоторые наследственные заболевания, которые повышают вероятность возникновения рака печени.
Цирроз. Болезнь, развивающаяся в результате формирования в печени рубцовой ткани и часто приводящая к раку. Наиболее существенные причины цирроза – употребление алкоголя и заболевание гепатитом С и В. Другая причина – это накопление в печени избыточного количества железа.
Употребление табака. Доказана связь между курением и возникновением рака печени. При сопутствующем употреблении алкоголя риск увеличивается.
Афлатоксины. Употребление продуктов, которые из-за неправильного хранения поражены афлотоксином В1(митотоксин гриба Аspergilis flavus), повышает риск заболевания. К таким продуктам относятся: пшеница, рис, кукуруза, соевые бобы, земляные орехи и пр.
Анаболические стероиды – это мужские гормоны, которые иногда используют спортсмены. Их длительное применение может несколько увеличить риск развития злокачественной опухоли печени.
Мышьяк. Есть страны, где употребляется вода, загрязненная мышьяком, что повышает риск возникновения рака печени.
Профилактика
Основными мерами профилактики рака печени являются:
- своевременная вакцинация от гепатита В;
- своевременное и качественное лечение гепатита В и С;
- лечение алкоголизма и полный отказ от алкоголя;
- регулярное наблюдение у гепатолога для больных циррозом и хроническими вирусными гепатитами (не реже 2-3 раз в год).
Особое значение имеет борьба с алкоголизмом, поскольку цирроз печени (особенно крупноузловая форма) обнаруживают примерно у 60-90% больных с гепатомой.
Вопросы и ответы
В разделе публикуются вопросы пациентов и ответы наших специалистов. Вопрос каждого человека касается конкретной проблемы, связанной с его заболеванием. Пациентам отвечают израильские клинические онкологи и главный врач LISOD, д.м.н., профессор Алла Винницкая.
Ответы специалистов основаны на знаниях принципов доказательной медицины и профессиональном опыте. Ответы соответствуют исключительно предоставленным сведениям, имеют ознакомительный характер и не являются врачебной рекомендацией.
Основная цель раздела – дать информацию пациенту и его семье, чтобы вместе с лечащим врачом принять решение о виде лечения. Предложенная Вам тактика лечения может отличаться от принципов, изложенных в ответах наших специалистов. Не стесняйтесь задать лечащему врачу вопрос о причинах отличий. Вы должны быть уверены, что получаете правильное лечение.
Здравствуйте. У моего отца поставили диагноз — обьемное оброзование s4 печени c инвазией стенки желчного пузыря, механическая желтуха. Скажите, доктор можно ли ему помочь, пробу не берут — снижают желтизну какая опухоль не знают. Помогите, пожалуйста, что делать. Спасибо.
Без гистологической верификации диагноза (биопсии) ничего определенного сказать нельзя. Это может быть первичная опухоль печени (печеночноклеточная или из желчных протоков), может быть вторичная опухоль (метастаз) из другого источника, или другая, более «экзотическая» опухоль. Параллельно следует сделать стадирование (определить распространенность процесса). После того как у нас будет гистологический диагноз и понятие о том местный ли это процесс или распространенный (метастатический) можно будет выстраивать план лечения.
Здравствуйте! В ходе УЗИ обнаружено образование в печени (около 1,9 см), предварительный диагноз — гепатома. Подскажите, какой метод диагностики применить, чтобы подтвердить или опровергнуть онкологию? Заранее благодарна за ответ!
Для начала необходимо сделать компьютерную томографию (КТ) или магнитно-резонансную томографию (МРТ) печени.
Здравствуйте! У моего близкого друга на КТ случайно обнаружили очаговое образование в печени. Мы в испуге, не знаем куда нам обращаться. Образование маленького размера, видно в одной проекции. Заранее благодарна
Если есть сомнения по поводу природы образования и оно больше одного см,то лучше всего — проведение ПЭТ-КТ исследования.
Добрый день. С наступающим Рождеством! Вопрос касается мамы. Был диагноз: Гемангиома печени. Расшифруйте диагноз пожалуйста. Какое лечение необходимо в данном случае? Возраст: 62 Жалобы пациента на данный момент: Боли Нет, присутствие органа имеется. Данные биопсии или гистологического исследования опухоли: нет Предшествующее лечение по поводу онкологического заболевания: нет Какое обследование уже было проведено: В динамике от 19.03.2010г., при СКТ печень обычной плотности-65 НY, однородной структуры,размерами -190/114/125 мл. В правой доле (с-7) сохраняется образование прежних размеров, до 20 мл в диаметре, с неровными контурами, слабо накапливающее контрастный препарат. Внутри печеночные желчные протоки не расширены . Желчный пузырь удален. Селезенка обычно расположена, плотность ее не изменена, в ее паренхиме множественные кальцинаты. Поджелудочная железа не увеличена в объеме , однородной структуре. Ретропанкреатическое пространство свободно. Объемных образований не выявлено. Параортальные лимфоузлы не увеличены надпочечники обычной формы и размеров. Почки без структурных изменений. В теле Тh21 позвонка нельзя исключить остеосклеротический очаг до 5 мл. ЗАКЛЮЧЕНИЕ: КТ- признаки очагового образования печени, более всего соответствует С-r, кальцинатов селезенки, с остеосклеротическим мтс тело Тh 11 позвонка, без отрицательной динамики от 19,03,10 г. Ответ буду ждать с нетерпением. С уважением к Вам Людмила.
Дорогая Людмила! Гемангиома — это доброкачественное образование из кровеносных сосудов. Оно не требует лечения при расположении в печени. Для дифференциальной диагностики от опухоли возможно проведение МРТ печени или сканирования с меченными радиоизотопом эритроцитами. Судя по тому что вы описываете (образования в печени и Th21 без динамики с марта 2010) речь не идет об онкологическом заболевании. Находка в 11 грудном позвонке может быть простым так называемым «костным островком».
У моего мужа месяц назад обнаружили на Узи и КТ множественные метастазы в печени от 12 до 45 мм, а сама опухоль не найдена нигде. Эти метастазы похожи на экране УЗИ на тёмные пятна. Ему 47 лет. Лечить его не стали, сказав мне, что в такой стадии рак уже не операбелен и неизлечим. Я ему про рак пока не говорю. Сейчас он очень слабый, потому что сильно худой. Вес 65 кг. Ходит медленно, иногда теряет равновесие, но ест всё, хотя и понемногу. Курит и бросать не собирается. Иногда жалуется на боли в области пупка и печени. Бывают дни, когда температура повышается до 37,5 и выше, и его начинает бить страшный озноб. Он становится очень холодный, губы синеют, начинается сильное потоотделение (как будто водой его окатили). Делаем ему тогда систему, в которую входит физраствор, глюкоза, преднизалон, спазмалгон и реосорбилакт. После системы потоотделение сначала усиливается, а потом прекращается и ситуация более-менее стабилизируется на некоторое время. Скажите мне, пожалуйста, это действительно рак? Или возможна ошибка. Можно что-то сделать, чтобы его вылечить или надо просто ждать конца, а все средства только на короткий срок отодвинут этот конец?
Юлия, добрый день! Ситуация не совсем понятна.
В любом случае необходимо провести стадирование и обязательно биопсию из очагов в печени для верификации диагноза. Только после этого будет понятно о каком заболевании и, соответственно, лечении будет идти речь.
Как алкоголь и сигареты влияют на организм человека
Употребление спиртных напитков и табакокурение объединяет то, что они вызывают привыкание. Причины, по которым человек выпивает свой первый стакан вина или выкуривает первую сигарету, как правило, одинаковые:
- Интерес к новым ощущениям;
- Желание не отстать от других, казаться взрослее;
- Влияние более опытных товарищей;
- Пример родителей, старших братьев.
Никто не собирается сознательно спиваться или доводить себя сигаретами до туберкулеза. Люди обычно не слушают предупреждений, уверены, что их беда коснуться не может, они бросят пить или курить тогда, когда им это надоест. Но привыкание к сигаретам и спиртным напиткам — это процесс физиологический, он от человека не зависит.
На свое пристрастие люди совершенно не жалеют денег и тратят астрономические суммы, лишь бы получить новую порцию алкоголя и выкурить сигарету. Только кроме денег, человек портит свое здоровье и медленно убивает себя. Конечно, может показаться, что если выкурить несколько волшебных никотиновых палочек и запить их сто граммами хорошего коньяка, никому ничего плохого не будет. Возможно, даже если визуально не видно, что человек пьян, организм тем временем уже испытывает дистресс. Курение и алкоголь оказывают свое негативное воздействие на все внутренние органы. К примеру, в первую очередь страдает сердце. Поэтому чаще всего алкоголики умирают от сердечных приступов, инсультов или инфарктов. Далее, страдают: печень, почки, органы пищеварения, легкие, желудок, головной мозг. Никотин и этанол вызывают физическую и психологическую зависимость.
У заядлого курильщика больше шансов заболеть раком легких, а у алкоголика – циррозом печени. Сначала человек и не подозревает, насколько вредны его пагубные привычки, но со временем у него ухудшается здоровье. Появляется одышка, повышается артериальное давление, учащается сердечный ритм. Более того, зависимые от табака и алкоголя, чаще страдают бессонницей, перепадами настроения, у них слабеет иммунная система.
Вред от сигарет
Как известно, в сигаретах содержится никотин, но кроме этого, в них много и других вредных веществ. Например, аммиак, мышьяк, смолы, формальдегид и прочие. Они самым отрицательным образом сказываются на здоровье человека. И если люди не видят то, что творится у них внутри, визуально всегда можно отличить курильщика от некурящего человека.
У некурящих людей свежий вид, бодрая походка. Курильщика же выдает серый, тусклый цвет лица, кожа становится дряблой. Он не может быстро пройти на длительную дистанцию, так как у него будет одышка. Таким людям тяжело заниматься спортом и вести здоровый образ жизни. Кроме всего прочего, у курящего человека страдают органы дыхания. При длительном курительном стаже у человека развиваются следующие заболевания: хронический бронхит, астма, трахеит, ларингит, рак горла или легких, воспаление легких и так далее. Также у курильщика начинают желтеть и портятся зубы, появляется плохой запах изо рта, меняются вкусовые качества, у мужчин появляется импотенция. Курению необходимо сказать твердое «нет».
Вред от алкоголя
Спиртосодержащие напитки настолько пагубно влияют на организм, что за некоторое время из здорового человека можно запросто превратиться в инвалида. Алкоголику очень тяжело избавиться от своей привычки, потому, что от алкоголизма страдает нервная система и зависимый медленно, но верно начинает деградировать. Вред для здоровья от алкогольных напитков заключается в том, что в них содержится спирт. Этанол после попадания в организм, мгновенно проникает в кровеносную систему, тем самым, поражая абсолютно все внутренние органы человека. Больше всего страдает сердце, печень, мозг и желудок, а это жизненно важные органы. Алкоголь способствует слипанию кровяных клеток, а значит, могут образоваться тромбы, закупорка вен и сосудов. Нет здоровых людей, которые употребляют алкоголь, пусть даже это хороший напиток. Со временем, все равно последствия дадут о себе знать. Не стоит забывать и о том, что этанол раздражает слизистую желудка, отсюда появляются такие заболевания желудочно-кишечного тракта, как: язва, гастрит. Но сильнейшее влияние спирт оказывает на головной мозг, он убивает клетки мозга, тем самым, нарушая его функционирование. После спиртных напитков человек медленно, но верно начинает деградировать. Его не волнуют проблемы, кроме одной – где взять очередную дозу. Он бросает работу, становится асоциальным и спивается. В итоге спиртосодержащие напитки могут привести к инвалидности, коме или летальному исходу.
Зачастую большинство алкоголиков являются курильщиками. Сочетая вместе эти вредные привычки, человек тем самым наносит здоровью двойной удар. Алкоголь и курение негативно влияют на центральную нервную систему. Люди становятся забывчивыми, так как у них ухудшается память, они перестают логически мыслить. Агрессия и раздражение — первый признак того, что зависимому жизненно необходима новая порция табака и этанола. Так как вред от алкоголя и сигарет оказывает токсическое действие, длительное употребление приводит к психическим расстройствам. Наиболее известное из них: алкогольный делирий, простыми словами – белая горячка. Кроме того, влияние на психику может проявляться следующим образом: человек становится рассеянным, быстро устает, возрастает чувство тревожности. Снижается трудоспособность, умственная активность, в некоторых случаях человек теряет чувство реальности и возвращается в нее, только покурив и выпив алкоголя.
Влияние курения и алкоголя на организм человека носит физический и психологический характер. Если говорить о психологии, то человек употребляет токсические вещества (алкоголь, табак, наркотики) по ряду причин.
Первой причиной можно считать снятие стресса. Так, если у человека неприятности или проблемы, он думает, что если пить и курить, то можно на время забыть обо всем. Возможно это и так, если сильно напиться и не соображать ни о чем. Но в результате, протрезвев, человек понимает, что проблемы не решены, а здоровье заметно подпорчено.
Вторая причина — средство привлечения внимания. Когда человек выпьет, он становится раскованнее, чувствует себя увереннее, а значит, может в баре или любом другом месте подойти к незнакомому человеку и познакомиться. Считает, что сигаретка придаст ему элегантности. Или, например, сидя в незнакомой компании, выпив, человек тянется за сигареткой, чтобы привлечь внимание и попросить зажигалку. На самом деле ситуаций много, но причина одна.
Третья причина — одиночество. Человеку грустно, он не знает с кем скоротать вечер, чем заняться, куда сходить, у него нет друзей. Вот и приходится ему выпивать под сигаретку с грустным видом, надеясь, что настроение поднимется. Но вместо этого на утро у него появляется похмельный синдром. И если к курению привыкание возникает не сразу, то алкогольные напитки быстро провоцируют зависимость. Надо сказать, что бросить курить сложно, но легче, чем сказать «нет» алкоголю.
Как избавиться от вредных привычек
Очень многие зависимые люди, которые признали свой недуг и «горят» желанием излечиться, возлагают большие надежды на кодировки. Всяческие блокаторы от зависимостей, не что иное, как психологический метод запугивания. Самый оптимальный способ избавиться от алкоголизма и табакокурения – сила воли. Если человек понял, что он болен и изъявил желание лечиться, это уже очень хороший знак. Теперь главное — держаться и верить в свои силы. Следует найти себе хобби, не думать о плохом, подобрать заменители токсических веществ. Возможно, если захочется выпить, лучше приготовить фруктовый коктейль или энергетический смузи. Сигареты заменить леденцами или орехами. И поменьше оставлять себе свободного времени. Самое главное — никогда не забывать, какой вред оказывают сигареты и алкоголь на здоровье. При желании выпить, нужно вспоминать, как плохо будет утром, что из-за алкоголя пострадает здоровье, и ухудшатся отношения в семье.
Побольше мотивации и веры в то, что нет ничего невозможного!
исследователей превращают клетки кожи в стволовые | Наука
Ученым удалось перепрограммировать клетки кожи человека непосредственно в клетки, которые выглядят и действуют как эмбриональные стволовые (ЭС) клетки. Этот метод позволяет генерировать стволовые клетки для конкретного пациента для изучения или лечения заболеваний без использования эмбрионов или ооцитов — и, следовательно, может обойти этические дебаты, которые преследуют эту область. «Это похоже на землетрясение как для науки, так и для политики исследования стволовых клеток», — говорит Джесси Рейнольдс, политический аналитик Центра генетики и общества в Окленде, штат Калифорния.
Работа основана на исследовании, опубликованном в прошлом году Шинья Яманака из Киотского университета в Японии, которое показало, что клетки хвоста мыши можно трансформировать в ES-подобные клетки путем вставки четырех генов ( Science NOW, 3 июля 2006 г.). Эти гены обычно отключаются после того, как эмбриональные клетки дифференцируются в различные типы клеток. В июне этого года Яманака и другая группа сообщили, что клетки действительно плюрипотентны, а это означает, что у них есть потенциал для роста в любой ткани тела ( Science NOW, 6 июня).
Теперь гонка за повторение подвига на человеческих клетках закончилась ничьей: сегодня две группы сообщают, что они перепрограммировали клетки кожи человека в так называемые индуцированные плюрипотентные клетки (iPC). В статье, опубликованной в Интернете в Cell , Яманака и его коллеги показывают, что их мышиный метод работает и с человеческими клетками. А в статье, опубликованной в Интернете в Science , Джеймс Томсон из Университета Висконсина, Мэдисон, и его коллеги сообщают об успехе в перепрограммировании человеческих клеток, опять же путем вставки всего четырех генов, два из которых отличаются от тех, которые использует Яманака.
В новой работе Яманака и его коллеги использовали ретровирус для переноса во взрослые клетки тех же четырех генов, которые они ранее использовали для репрограммирования клеток мыши: OCT3 / 4 , SOX2 , KLF4 и c-MYC. . Они перепрограммировали клетки кожи лица 36-летней женщины и соединительной ткани 69-летнего мужчины. Примерно одна клеточная линия iPC была произведена на каждые 5000 клеток, обработанных исследователями с помощью этого метода, эффективность, которая позволила им получить несколько клеточных линий из каждого эксперимента.
Команда Томсона начала с нуля, определив собственный список из 14 генов-кандидатов на перепрограммирование. Как и группа Яманаки, команда использовала систематический процесс исключения, чтобы идентифицировать четыре фактора: OCT3 и SOX2 , как использовал Яманака, и два разных гена, NANOG и LIN28 . Группа перепрограммировала клетки кожи плода и крайней плоти новорожденного мальчика. По словам Томсона, исследователям удалось трансформировать примерно одну из 10 000 клеток, что меньше, чем достигла методика Яманаки, но все же достаточно, чтобы создать несколько клеточных линий в одном эксперименте.
Несмотря на то, что оба метода многообещающие, у них есть и обратная сторона. Ретровирусы, используемые для вставки генов, могут вызывать опухоли в тканях, выросших из клеток. Все согласны с тем, что следующий решающий шаг — найти способ перепрограммировать клетки путем включения генов, а не вставки новых копий. По словам исследователя стволовых клеток Дугласа Мелтона из Гарвардского университета, эта область быстро движется к этой цели. «Нетрудно представить время, когда можно было бы добавить небольшие молекулы, которые будут щекотать те же сети, что и эти гены», и произвести перепрограммированные клетки без генетических изменений, — говорит он.
Как только недостатки будут устранены, «вся эта область полностью изменится», — говорит исследователь стволовых клеток Хосе Сибелли из Университета штата Мичиган в Ист-Лансинге. «Людям, занимающимся этикой, придется найти что-то новое, о чем можно будет беспокоиться».
Более подробные новости по этой теме можно найти в выпуске Science за эту неделю, который доступен онлайн 22 ноября.
Связанный сайт
Архей на коже человека
Abstract
Недавняя эра изучения микробиома человека предоставила ценную информацию о микробных обитателях, полезных и патогенных микроорганизмах.Усилия по скринингу, основанные на секвенировании ДНК, выявили тысячи бактериальных линий, связанных с кожей человека, но предоставили лишь неполную и грубую информацию об архее. Здесь мы впервые сообщаем о количественной оценке и визуализации архей на коже человека. На основе 16 копий гена S рРНК археи составляли до 4,2% микробиома кожи прокариот. Большинство проанализированных генных сигнатур принадлежали Thaumarchaeota, группе архей, которых мы также обнаружили в больницах и чистых комнатах.Метаболический потенциал окисления аммиака у связанных с кожей архей подтвержден успешным обнаружением таумархейных генов amoA в образцах кожи человека. Однако активность и возможное взаимодействие с человеческими эпителиальными клетками этих ассоциированных архей остается открытым вопросом. Тем не менее, в этом исследовании мы приводим доказательства того, что археи являются частью микробиома кожи человека, и обсуждаем их потенциал обмена аммиака на коже человека.
Образец цитирования: Probst AJ, Auerbach AK, Moissl-Eichinger C (2013) Археи на коже человека.PLoS ONE 8 (6): e65388. https://doi.org/10.1371/journal.pone.0065388
Редактор: Джош Нойфельд, Университет Ватерлоо, Канада
Поступила: 11 февраля 2013 г .; Принята к печати: 29 апреля 2013 г .; Опубликован: 12 июня 2013 г.
Авторские права: © 2013 Probst et al. Это статья в открытом доступе, распространяемая в соответствии с условиями лицензии Creative Commons Attribution License, которая разрешает неограниченное использование, распространение и воспроизведение на любом носителе при условии указания автора и источника.
Финансирование: Финансирование предоставлено Институтом микробиологии и Центром архей (Майкл Томм), Регенсбургским университетом и Европейским космическим агентством (контракт № 4000103794/11 / NL / PA по субподряду с DLR (D / 316 / 67130713)) подтверждается. AJP был поддержан Национальным немецким академическим фондом (Studienstiftung des deutschen Volkes). Финансирующие организации не играли никакой роли в дизайне исследования, сборе и анализе данных, принятии решения о публикации или подготовке рукописи.
Конкурирующие интересы: Авторы заявили об отсутствии конкурирующих интересов.
Введение
Археи долгое время считались древней формой микроорганизмов, обитающих в экстремальных условиях. Однако картина архей изменилась за последнее десятилетие, когда эти организмы были обнаружены в большом количестве в холодных и умеренных средах по всему миру [1].
Археи также могут играть важную роль в организме человека, поскольку метаногенные археи могут составлять до 12% от общего количества анаэробов в кишечнике человека [2].В полости рта метаногены связаны с некоторыми заболеваниями пародонта [2], хотя патогенез архей еще не подтвержден.
Проект «Микробиом человека», основанный для расшифровки всего набора микроорганизмов, связанных с человеческим телом, продолжает предоставлять ценную информацию о том, как микробное разнообразие соотносится с состоянием здоровья человека [3]. К настоящему времени бактериальная динамика крупнейшего человеческого органа, кожи, подробно изучена [4], в то время как только два исследования сообщают об обнаружении архей на коже человека [5], [6].Халкр и его сотрудники изучили 60 пупков и обнаружили три различных филотипа архей, незначительно появляющихся в большом подмножестве бактериальных последовательностей. Выявленные филотипы архей принадлежали к Euryarchaeota и были извлечены только из 6 образцов. Авторы обнаружили галофилы ( Halobacteriaceae ) в двух образцах и метаногены ( Methanobrevibacter ) в пяти различных образцах. Два из трех филотипов архей были получены от человека, который годами не принимал душ, что означает, что археи составляют лишь незначительную часть микробиома пупка и кожи.Более того, обнаруженные таксоны архей ранее были обнаружены связанными с ротовой полостью человека и желудочно-кишечным трактом человека и, таким образом, вероятно, являются оральными или фекальными контаминантами [7], [8]. Хотя Халкр и его сотрудники утверждали, что первыми сообщили об архее в микробиоме кожи человека, Caporaso et al. [6] уже сообщал о подписях архей, в частности Thaumarchaeota, в образцах, взятых с ладоней двух особей. Однако они определили эти микроорганизмы как второстепенную, временную часть человеческого микробиома и взяли на себя незначительную роль.
Тем не менее, оба исследования обнаружили архей посредством совместной амплификации их генов 16 S рРНК вместе с бактериями. Это, а также тот факт, что использованные пары праймеров не полностью соответствовали (таум-) генам 16 S рРНК архей, не позволяет сделать вывод о роли, численности или разнообразии архей на коже человека. Например, пара праймеров F515 / R806 (использованная в [6]) без несоответствия поражает только 50% всех архей и, в частности, 8% всех таумархей. Совершенного совпадения в почвенной кренархеотической группе (I.1б), который включает Candidatus Nitrososphaera (SILVA TestPrime [9]).
Исследования микробиомов в антропогенных средах, таких как чистые комнаты, которые находятся под сильным влиянием человеческого (кожного) микробиома, показали, что археи постоянно присутствуют [10], [11]. Большинство из этих архей были Thaumarchaeota, недавно предложенным типом, включающим назначенные окислители аммиака [12]. В связи с недавней переклассификацией типа таумархей и, следовательно, отнесением определенных групп кренархей к кладе таумархей, недавнее исследование LaDuc и соавторов [13] ошибочно заявило о первых доказательствах существования таумархей в условиях чистых комнат, хотя эта группа были идентифицированы ранее [10], [11].До сих пор эти чистые комнатные археи принадлежали в основном к таумархеальной кладе I.1b, представители которой обычно встречаются в почвенном микробиоме, где они, вероятно, вносят вклад в глобальный цикл азота [14]. Вероятная связь с людьми обсуждалась Мойсл-Эйхингером [11], но вопрос о том, происходят ли эти археи из кожи человека, остался без ответа.
В настоящем исследовании мы решаем вопрос, может ли кожа человека быть носителем или даже средой обитания архей. Мы показываем, что археи и, в частности, таумархеи представляют собой обнаруживаемую часть микробиома кожи человека, и их сигнатуры тесно связаны с сигнатурами, обнаруженными в больницах и чистых комнатах.Кроме того, мы предоставляем информацию о морфологии клеток и функциональных генах архей на коже человека.
Материалы и методы
Образцы кожи человека были взяты и обработаны с одобрения и в соответствии с Комиссией по этике Университета Регенсбурга. Комиссия по этике заявила, что применяемые методы не вызывают никаких этических проблем, и утвердила следующие процедуры. Устное информированное согласие было получено от всех участников исследования, что согласуется с заявлением Комиссии по этике.Каждый участник сдал образец сразу после самостоятельной выборки, и устное согласие документировалось вручную вместе с получением образцов. Образцы обрабатывались анонимно. Человеческий материал не был предметом этого исследования. Образцы микробов или полученные данные не могут быть приписаны определенному человеку.
Образцы со всей передней части туловища были взяты самими добровольцами с помощью салфеток, не содержащих ДНК. Испытуемым было предложено тщательно вытереть туловище (держа салфетку без ДНК стерильной перчаткой) перед тем, как принять обычный душ.Перед взятием проб добровольцы не применяли косметику. Салфетки немедленно хранили на льду или замораживали перед обработкой. Обзор всех образцов салфеток из кожи человека приведен в таблице S1.
Отбор проб из окружающей среды в помещении проводился либо с помощью наборов для отбора биологических проб (BiSKit, QuickSilver Analytics, Abingdon, MD, USA, в соответствии с инструкциями производителя), либо с помощью предварительно увлажненной салфетки без ДНК, прикрепленной к не содержащей ДНК. инструмент для отбора проб из стали (Таблица S1).Был взят образец из одного комплекса чистых помещений (EADS, Фридрихсхафен, Германия) с чистыми помещениями ISO 5 и ISO 8. Чистые помещения находились в полностью сертифицированных рабочих условиях. Дополнительными точками отбора проб были два отделения интенсивной терапии, одно в Регенсбурге (Германия) и одно в Граце (Австрия), оба постоянно работали (Таблица S1).
Экстракция образца из салфеток (включая встряхивание и обработку ультразвуком) выполнялась в 40 мл воды для ПЦР (для молекулярных анализов) или фосфатно-солевого буфера (PBS-буфер, для флуоресценции in situ гибридизация), как описано в другом месте [ 11].Перед обработкой жидкости концентрировали до 200–500 мкл с использованием фильтровальных трубок Amicon 50 (Millipore, Billerica, MA, USA).
Эффективность восстановления устройств для отбора проб сильно зависит от инструмента для отбора проб и пористости поверхности [15], [16]. Было доказано, что отбор проб с непористых поверхностей обычно имеет низкую эффективность восстановления — 8–20% [16], [17]. Тот факт, что кожа человека представляет собой очень пористую поверхность, и отбор образцов кожи проводился неспециалистами (самостоятельный отбор образцов) в сочетании с низкой эффективностью восстановления образцов в целом, позволяет сделать вывод о том, что была подвергнута только небольшая часть микробиома кожи. выздоровел.Однако ожидается, что соотношение извлеченных бактерий и архей не будет зависеть от эффективности отбора проб.
Обработка моноазидом пропидия на выбранных образцах из отделения интенсивной терапии в Регенсбурге была выполнена, как описано в другом месте [18], до экстракции ДНК. Моноазид пропидия (PMA) представляет собой химическое вещество, которое внедряется в доступные молекулы ДНК в данном растворе и образует ковалентную связь после фотоактивации азида. После связывания PMA ДНК маскируется и больше не доступна для амплификации с помощью ПЦР.Клетки с интактными клеточными мембранами не проникают PMA, их ДНК остается немеченной для амплификации ПЦР и, следовательно, может быть обнаружена. Этот анализ позволяет различать микробное сообщество с нарушенной мембраной и жизнеспособное микробное сообщество.
Экстракция ДНК была выполнена комбинацией взбивания шариков и метода XS-буфера, описанного в Moissl-Eichinger [11], протоколе, адаптированном для сред с низким содержанием биомассы. Перед нанесением XS-буфера было включено взбивание шариков, чтобы гарантировать, что также лизируются выносливые микробы (пробирки для взбивания шариков были взяты из набора для выделения ДНК MO BIO Power Biofilm ™, MO BIO, Карлсбад, Калифорния, США).После взбивания шарики промывали 400 мкл чистой воды, пригодной для ПЦР, для уменьшения потерь образца.
Количественная ПЦР бактериальных и архейных последовательностей гена 16 S рРНК была проведена в трех повторностях, как описано в другом месте, с парами праймеров 338 bf / 517 ur и 344 af / 517 ur соответственно [11], [19] (конечная концентрация праймера : 300 нМ). 16 генов S рРНК из геномной ДНК архейных и бактериальных эталонных штаммов Methanosarcina barkeri и Bacillus safensis были амплифицированы с наборами праймеров 8 af / 1406 ur и 9 bf / 1406 ur [20], [21].Количественную оценку стандартов проводили с помощью Qubit Quantitation Platform 2.0 (High Sensitivity Kit, Invitrogen, Carlsbad, CA, USA). Было проведено сорок циклов кПЦР (Quantitect SYBR Green PCR Mix, Qiagen, Hilden, Германия) (система RotorGene 6000 Real-Time PCR, Corbett Life Science, Concorde, NWS, Австралия) с начальной денатурацией при 95 ° C в течение 15 минут и протокол циклирования следующий: денатурация при 94 ° C в течение 15 секунд, отжиг при 60 ° C в течение 30 секунд и удлинение при 72 ° C в течение 30 секунд. Кривая плавления построена при 72–95 ° C.Эффективность КПЦР находилась в диапазоне от 0,87 до 0,92, а значения R 2 стандартных кривых находились в диапазоне от 0,98 до 1,00. Пределы обнаружения были определены как 105 копий / мкл для количественной ПЦР архей (порог для контрольных образцов без матрицы) и 334 копий / мкл для количественной ПЦР бактерий. Эти пороговые значения были получены следующим образом: qPCR для отрицательного контроля архей выявила минорные сигналы от димеризации праймера, что было подтверждено с помощью гель-электрофореза. Чтобы исключить такой фон, отрицательные контроли qPCR усредняли и использовали в качестве порогового значения.Сигнал от архей не был получен от контрольных образцов, не являющихся матрицами, и экстрактов. Поскольку реагенты для количественной ПЦР не были свободны от бактериальной ДНК, как сообщалось в другом месте [22], предел обнаружения для бактериальной КПЦР был соответственно увеличен (334 копий / мкл, усредненная бактериальная КПЦР без контроля матрицы).
Амплификацию генов 16 S рРНК архей из образцов проводили либо напрямую (пара праймеров 344 af / 915 ar; [23], [24]), либо с помощью вложенной ПЦР (пары праймеров 8 af / UA1406 R [25] , [26] и 340 af / 915 ar [27], [23]; 2 × 35 циклов).Покрытие праймеров оценивали с помощью TestProbe, инструмента сопоставления и оценки зонда SILVA по всей базе данных 108 SILVA SSU Ref [28], которая выявила те комбинации праймеров с наибольшим охватом по сравнению с другими изученными комбинациями. Тестируемые комбинации включали следующие праймеры. Нападающие: 8af [20], 27FLP [29], 21af [30], 340af [27], 344af [31], A751f [26]. Реверс: 909r [32], 915r [24], 1000R [27], 1100R [33], 1119ar [34], UA1406r [26], 1391r [32], 1406ur [21], 1492ur [21]).
Клонирование продуктов ПЦР гена 16 S рРНК, секвенирование и анализ выполняли следующим образом: продукты ПЦР клонировали в компетентных клетках TOPO10 (Invitrogen, TOPO® TA Cloning® Kit, pCR®2.1), вставки положительных клонов подвергали скринингу с использованием двух ( Hinf I; Bsu RI) и четырех ( Alu I; Hha I; Hinf I; Rsa ) рестрикционных ферментов после ПЦР-амплификации. с указанными выше парами праймеров. Клоны, несущие уникальные вставки (52 уникальных образца), секвенировали по Сэнгеру (секвенировали 100 вставок), обрезали, проверяли качество (уничтожитель химер), выравнивали [28] и группировали с уровнем разницы в 1% с помощью программного пакета mothur [35] и отнесены к ним. в качестве операционных таксономических единиц (OTU) в рукописи.Затем репрезентативные последовательности из каждой OTU были классифицированы с использованием байесовского классификатора [35] по сравнению с вручную созданной базой данных GreenGenes, которая содержит представителей 98% идентичных кластеров и обновленную таксономию [36] (доступна по адресу http://www.secondgenome.com/go / 2011-greengenes-taxonomy /). Полученные последовательности гена 16 S рРНК архей были представлены в GenBank (№ JX865653-JX865767).
Филогенетическое дерево репрезентативных последовательностей OTU (выровненных по SINA; 28) было вычислено с использованием ARB [37] и версии базы данных SILVA SSU111 [38] с применением алгоритма максимального правдоподобия.Последовательности были обрезаны до одинаковой длины выравнивания перед расчетом дерева ( E. coli , позиция 346–943). Топология дерева подтвердила классификацию, полученную с помощью байесовского метода (см. Выше).
Обнаружение генов amoA выполняли, как описано в Tourna et al. , 2011 [39] с использованием праймеров amoA104F-1d (GCA GGA GAC TAC ATM TTC TA) и amoA616R (GCC ATC CAT CTG TAT GTC CA). Клонировали ампликоны и секвенировали по Сэнгеру с использованием праймеров M13.Объединенный образец всех людей, исследованных в этом исследовании, использовали в качестве матрицы для ПЦР (5 мкл на 20 мкл реакции ПЦР). Было получено 32 последовательности, 28 были высокого качества (хорошее качество хроматограммы,> 800 п.н.) и 21 из них было идентифицировано как генов amoA с помощью BLAST [40] в базе данных NCBI (депонировано под номерами доступа KC582378-KC582398. ). Семь последовательностей (приблизительно 510 п.н.) показали высокое сходство с последовательностями из Staphylococcus epidermidis (предшественник липазы, gehD ) и не были включены в анализ. Гены AmoA были сгруппированы с 97% сходством с использованием mothur [35], а дерево максимального правдоподобия было вычислено в ARB с помощью базы данных amoA от Pester et al. , 2012 [37].
Контроли для молекулярного анализа для каждого отдельного шага (отбор образцов, экстракция ДНК, установка ПЦР) и по крайней мере один холостой контроль, который прошел все процедуры обнаружения, включая регулярную ПЦР архей, вложенную ПЦР, количественную ПЦР и бактериальную КПЦР. .Все контроли, направленные на архей, были отрицательными, видимых полос не наблюдалось ни при какой ПЦР-амплификации. Бланки BiSKit показали определенное количество детектируемых бактериальных генов 16 S рРНК (образцы из отделения интенсивной терапии Граца и чистой комнаты Фридрихсхафена; архейный контроль был отрицательным). Поэтому все образцы людей и образцы из отделения интенсивной терапии в Регенсбурге были взяты с помощью салфеток, не содержащих ДНК (сухая термообработка в течение 24 часов, 170 ° C). Все холостые экстракты протирания были отрицательными в каждой (q) ПЦР архей и бактерий.
Флуоресцентная гибридизация in situ (FISH) была проведена на одном образце кожи человека. Образец фиксировали в параформальдегиде (3% (мас. / Об.), Конечная концентрация). FISH проводили, как описано ранее [41], с использованием зондов ARC915 (направленных против архей, помеченных родаминовым зеленым) и EUB338 / I (направленных против бактерий, меченных CY3), 20% (об. / Об.) Формамида и 0,01% (мас. / Об. ) SDS. DAPI использовался в качестве контрастного красителя. NONEUB-Probe использовался в качестве бессмысленного отрицательного контроля. Микроскопию выполняли с использованием микроскопа Olympus BX53 (Olympus, Гамбург, Германия; камера: Olympus XM10, программное обеспечение: CelSens Standard 1.5). Флуоресцентную микроскопию выполняли с использованием следующих фильтров. U-FUN (возбуждение 360–370 нм, излучение 420 нм IF) для DAPI, U-MINB3 (возбуждение 470–495 нм, 510 нм IF) для родамина зеленого и U-FRFP (возбуждение 535–555 нм HQ, излучение 570 –625 HQ) для CY3. В образцах FISH обычно обнаруживается большое количество частиц (также из салфетки, см. Процедуру экстракции). Эти частицы были либо флуоресцентно-активными с фильтром CY3, либо со всеми тремя фильтрами, четко отличимыми от микроорганизмов и не включались в анализ.Эти волокна сделали невозможным количественный анализ FISH.
Результаты и обсуждение
Для исследования численности и разнообразия архей на коже человека были взяты образцы салфеток от 13 человек (7 женщин и 6 мужчин, возраст от 20 до 40 лет, целые передние туловища, таблица S1) и проанализированы с помощью чувствительных молекулярных методов. У всех людей было обнаружено присутствие генов 16 S рРНК архей на коже, что составляет до 4,23% от всего восстановленного микробиома прокариот (0.В среднем 60%, рис. 1). Поскольку бактерии, вероятно, обладают большим количеством рибосомных генов на геном (в среднем 4,17) по сравнению с археями (1,69) [42], средняя доля архейных клеток может быть даже больше (1,40% в среднем, макс. 9,86%).
Рис. 1. Обилие бактериальных и архейных копий гена 16 S рРНК, извлеченных из передних торсов 13 человек.
Значения над гистограммами показывают процент копий архейных генов во всем обнаруженном прокариотическом микробиоме. Звездочки указывают на процент архей ниже 0.01. Ось X показывает номер образца человека (Таблица 1), ось Y показывает логарифмически трансформированные количества 16 генов S рРНК.
https://doi.org/10.1371/journal.pone.0065388.g001
Для более глубокого анализа разнообразия архей на коже человека были отобраны пять образцов. Структура сообщества архей состояла из OTU типа Thaumarchaeota (88% от всех OTU) и Euryarchaeota (12%) с 17 различными таксономическими OTU в общей сложности. Все люди демонстрировали последовательности Thaumarchaeota.Филогенетический анализ этих последовательностей кожи архей поместил их близко к археям, окисляющим аммиак из почвы (Thaumarchaota, группа I.1b, рис. 2), но, что интересно, также близко к последовательностям из искусственной среды (чистые комнаты, отделения интенсивной терапии), обнаруженным в этой и более ранние исследования [11] (рис. 2). Помимо сигнатур таумархей, один человек обнаружил две последовательности эвриархей, которые принадлежали таксону Methanosarcina , предполагаемому метаногену, о котором ранее сообщалось для сельскохозяйственных сред, но также и для кишечника человека [2].Классификация последовательностей сведена в Таблицу 1.
Рис. 2. Дерево максимального правдоподобия, отображающее все обнаруженные OTU на коже человека, в отделении интенсивной терапии и в чистых помещениях.
Символ « человек »: филотип, извлеченный из кожи человека (количество символов дает количество особей, несущих этот филотип; 5 субъектов были проверены на предмет генофонда архей 16 S рРНК). Обозначение « больница » (квадрат с крестом): филотип обнаружен в отделении интенсивной терапии (обследовано два отделения интенсивной терапии).Обозначение « квадрат »: обнаружено в чистом помещении для сборки космического корабля (проанализировано одно помещение). Символ « star » выделяет филотипы, которые также были обнаружены в образце, обработанном моноазидом пропидия (PMA), то есть в клетках с интактными мембранами. Масштабная линейка относится к 10% заменам нуклеотидов. В качестве внешней группы использовались Pyrobaculum arsenaticum и Thermofilum pendens (Crenarchaeota).
https://doi.org/10.1371/journal.pone.0065388.g002
Таблица 1. Анализ библиотек клонов и классификация таксономических OTU.
Присутствие архей на коже человека было дополнительно подтверждено флуоресцентными экспериментами по гибридизации in situ (рис. 3). К сожалению, относительное обилие архей по сравнению с бактериальными клетками невозможно было рассчитать из-за большого количества флуоресцентно-активных частиц и волокон в образцах (подробности см. В разделе «Материалы и методы»), но сигналы архей были очевидны и легко обнаруживались.Клетки архей визуализировались в микробиоме кожи как маленькие кокки (примерно 0,5 мкм в диаметре). Их форма и размер были подобны клеткам таумархей, ранее обнаруженным в образцах ила [43].
Рис. 3. Флуоресценция in situ гибридизация, выполненная на образце салфетки кожи человека для визуализации архей.
ДНК-содержащая клетка (окраска DAPI): синий, археи: зеленый, бактерии: красный. I-V: показаны примеры положительных сигналов архей (маленькие кокки, зонд ARC915, помеченный родаминовым зеленым), которые дают положительный сигнал с помощью DAPI и отсутствие сигнала с помощью зонда, направленного на бактерии (EUB338 / I, помеченного CY3).VI: Пример положительного бактериального сигнала. Полоса: 2 мкм.
https://doi.org/10.1371/journal.pone.0065388.g003
Поскольку среда, в которой доминирует человек, отражает микробное разнообразие, связанное с кожей и телом человека [44], теперь мы можем предложить логическую причину более раннего открытия. сигнатур архей в контролируемых чистых комнатах по всему миру [10], [11], [13]. Присутствие (таум-) архейных сигнатур в чистых помещениях было подтверждено в этом исследовании (объект EADS для чистых помещений) и в дальнейшем распространено на больницы.Были взяты образцы из двух отделений интенсивной терапии, и было подтверждено присутствие таумархеот в этих искусственных средах (рис. 2). До сих пор все антропогенные среды, изученные авторами, выявили присутствие генов 16 S рРНК архей, принадлежащих к двум разным типам, Thaumarchaeota и Euryarchaeota, что можно отнести к используемым чувствительным и подходящим методам. Целостность и, следовательно, вероятная жизнеспособность архейных клеток в образцах салфеток для пола также была подтверждена FISH [11] и молекулярным анализом жизнеспособности (таблица S1, рис.2). Эвриархеи чистых помещений включали метаногены и различные галофилы; обе группы были связаны с микрофлорой ротовой полости и кишечника человека [2], [8], а недавно были также обнаружены в пупке [5]. Однако вышеупомянутое исследование пупков не выявило каких-либо признаков таумархей и только три филотипа эвриархей в очень небольшой подгруппе образцов [6]. Euryarchaeota (снова метаногены и галофилы), Crenarchaeota и Thaumarchaeota были обнаружены в исследовании Caporaso et al., 2011, который проанализировал микробное сообщество кишечника, языка и ладоней человека только у двух человек, но в 396 временных точках. В частности, микробиом пальмы выявил (колеблющееся) присутствие родственных последовательностей Nitrososphaera . На основании своего статистического анализа авторы сочли эти организмы незначительными и временными членами микробиома человека. Безусловно, кожа ладони представляет собой одну из основных поверхностей контакта человека с его биотической и абиотической средой, и временные микроорганизмы можно найти там больше, чем где-либо еще на коже человека.Следовательно, мы исключили человеческие ладони и сосредоточились на коже туловища человека, которая может иметь более типичное, менее подверженное влиянию микробное разнообразие. Основываясь на результатах исследования 13 образцов кожи человека, мы утверждаем, что Caporaso et al. , возможно, недооценил важность архей и особенно таумархей на коже человека. Чтобы подчеркнуть обнаружение общего присутствия архей на коже человека (все образцы выявили признаки архей), мы даже смогли визуализировать клетки архей, что указывало на их активный физиологический статус и их очевидное присутствие в образцах с кожи человека.Мы можем сделать вывод, что предыдущие исследования либо (методически) упускали из виду разнообразие архей, связанных с кожей человека, или недооценивали их изобилие.
Наше исследование было первым, систематически показывающим, что археи, в частности Thaumarchaeota, постоянно присутствуют на коже человека. Это открытие приводит к ряду вопросов, на которые невозможно ответить при нынешнем уровне знаний. Например, роль, метаболизм, скорость заражения или также происхождение архей, связанных с кожей, на сегодняшний день неясны и требуют изучения в последующих исследованиях.
Интересно, что представители кластера Thaumarchaeota I.1b были обнаружены в почве и водной среде, а также на очистных сооружениях, как недавно сообщалось [43], [45]. Обнаружение таких таумархейных последовательностей и клеток на коже человека и в искусственной среде может указывать на новые, в настоящее время неизвестные роли и метаболические возможности помимо хемолитоавтотрофии [43]. Однако мы отмечаем, что все культивируемые до сих пор виды таумархей являются окислителями аммиака [12], [39], [46], [47], а кожа человека постоянно выделяет небольшое количество аммиака [48].Мы смогли амплифицировать и секвенировать генов amoA из объединенной выборки всех 13 человек. Анализы мегабластов против коллекции нуклеотидов NCBI четко показали, что эти последовательности принадлежат генам amoA (значение E = 0). Филогенетическое дерево извлеченных последовательностей amoA показано на рисунке 4. Полученные последовательности не были тесно связаны с генами Nitrososphaera viennensis или N. gargensis amoA , но были связаны с двумя подкластерами Nitrososphaera и , Инжир.4), как и большинство генов 16 S рРНК (рис. 2).
Рис. 4. Дерево максимального правдоподобия на основе последовательностей гена архей amoA .
Последовательности, восстановленные в этом исследовании, выделены жирным шрифтом. Информация в скобках указывает количество извлеченных последовательностей. Столбик означает 10% замен нуклеотидов на сайт.
https://doi.org/10.1371/journal.pone.0065388.g004
Это открытие предлагает по крайней мере одно возможное объяснение присутствия этих микроорганизмов.Можно предположить, что хемолитотрофный оборот аммиака у Thaumarchaeota может влиять на регуляцию pH кожи человека и, следовательно, на естественный защитный слой, но это еще предстоит доказать. Кроме того, взаимодействие человека с археями, по-видимому, не ограничивается пассивной и / или косвенной метаногенной активностью в толстой кишке и ротовой полости, но может быть активным и прямым взаимодействием. Поскольку протоколы, используемые для Проекта микробиома человека, очевидно, недооценивают присутствие архей в целом, методы скрининга в науке и медицине должны быть лучше ориентированы на улучшение обнаружения архей, что затем приведет к всестороннему пониманию их полезной или потенциально патогенной роли в микробиом человека (кожи).
Благодарности
Мы благодарим Роберта Хубера, Райнхарда Вирта, Эмму Гаген, Харальда Хубера, Шарифа Османа за ценные обсуждения и предложения. Мы благодарны командам из Фридрихсхафена (EADS Astrium), Граца (Габриэле Берг, Universitätsklinik Graz) и Регенсбурга (Caritas-Krankenhaus St. Josef) за возможность получить доступ и опробовать чистые помещения и отделения интенсивной терапии. Благодарим людей, предоставивших образцы кожи.
Вклад авторов
Задумал и спроектировал эксперименты: AJP CM.Проведены эксперименты: AJP AKA CM. Проанализированы данные: AJP CM. Предоставленные реагенты / материалы / инструменты анализа: AJP AKA CM. Написал статью: AJP CM.
Ссылки
- 1. Карнер М.Б., Делонг Е.Ф., Карл Д.М. (2001) Доминирование архей в мезопелагиали Тихого океана. Природа 25: 507–510.
- 2. De Macario EC, Macario AJL (2009) Метаногенные археи в здоровье и болезни: новая парадигма микробного патогенеза. Int J Med Microbiol 299: 99–108.
- 3. Консорциум проекта микробиома человека (2012) Структура, функции и разнообразие здорового микробиома человека. Природа 486: 207–214.
- 4. Грайс Э.А., Сегре Я.А. (2011) Микробиом кожи. Nat Rev Microbiol 9: 244–253.
- 5. Халкр Дж., Латимер А.М., Хенли Дж. Б., Раунтри Н. Р., Фирер Н. и др. (2012) Там джунгли: бактерии в пупке очень разнообразны, но предсказуемы. PLoS One 7 (11): e47712.
- 6. Капорасо Дж. Г., Лаубер С. Л., Костелло Е. К., Берг-Лайонс Д., Гонсалес А. и др.(2011) Движущиеся картинки микробиома человека. Геномная биология 12: R50.
- 7. Лепп П.У., Бриниг М.М., Оуверни С.К., Палм К., Армитаж Г.К. и др. (2004) Метаногенные археи и пародонтоз человека. Proc Natl Acad Sci USA 101: 6176–81.
- 8. Оксли А. П., Ланфранкони М. П., Вюрдеманн Д., Отт С., Шрайбер С. и др. (2010) Галофильные археи в слизистой оболочке кишечника человека. Environ Microbiol 12: 2398–2410.
- 9. Клиндворт А., Прюсс Э., Швир Т., Пеплис Дж., Кваст С. и др.(2012) Оценка общих праймеров для ПЦР гена 16 S рибосомной РНК для классических исследований разнообразия на основе секвенирования и исследований следующего поколения. Nucleic Acids Res. 41: e1
- 10. Moissl C, Bruckner JC, Venkateswaran K (2008) Анализ архейного разнообразия чистых помещений для сборки космических аппаратов. ISME J 2: 115–119.
- 11. Moissl-Eichinger C (2011) Археи в искусственной среде: их присутствие в глобальных чистых помещениях космических кораблей и влияние на планетарную защиту. ISME J 5: 209–219.
- 12. Пестер М., Шлепер С., Вагнер М. (2011) Таумархеи: новый взгляд на их филогению и экофизиологию. Curr Opin Microbiol 14: 300–306.
- 13. LaDuc MT, Vaishampayan P, Nilsson HR, Torok T, Venkateswaran K (2012) Разнообразие бактерий, архей и грибов, полученное в результате пиросеквенирования, в аппаратной части космического корабля, предназначенной для Марса. Appl Environ Microbiol 78: 5912–5922.
- 14. Лейнингер С., Урих Т., Шлотер М., Шварк Л., Ци Дж. И др.(2006) Среди прокариот, окисляющих аммиак, преобладают археи. Природа 442: 806–9.
- 15. Probst A, Facius R, Wirth R, Moissl-Eichinger C (2010) Валидация протокола нейлонового тампона для эффективного извлечения бактериальных спор с гладких и шероховатых поверхностей. Appl Environ Microbiol 76: 5148–5158.
- 16. Пробст А., Фасиус Р., Вирт Р., Вольф М., Мойсл-Эйхингер С. (2011) Восстановление контаминантов спор Bacillus с шероховатых поверхностей: проблема контроля чистоты космических полетов.Appl Environ Microbiol 77: 1628–1637.
- 17. Kwan K, Cooper M, LaDuc MT, Vaishampayan P, Stam C, et al. (2011) Оценка процедур сбора, обработки и анализа биомолекул с поверхностей с низким содержанием биомассы. Appl Environ Microbiol 77: 2943–2953.
- 18. Nocker A, Sossa-Fernandez P, Burr MD, Camper AK (2007) Использование моноазида пропидия для различения живых и мертвых в микробной экологии. Appl Environ Microbiol 73: 5111–5117.
- 19.Пробст AJ, Holman H-YN, DeSantis TZ, Andersen GL, Birarda G и др. (2012) Борьба с меньшинством: сульфатредуцирующие бактерии в подповерхностной биопленке с преобладанием архей. ISME J 7: 635–651.
- 20. Burggraf S, Olsen GJ, Stetter KO, Woese CR (1992) Филогенетический анализ Aquifex pyrophilus . Syst Appl Microbiol. 15: 352–356.
- 21. Lane DJ (1991) Секвенирование 16 S / 23 S рРНК. В: Stackebrandt E, Goodfellow M, редакторы. Методы нуклеиновых кислот в бактериальной систематике.Нью-Йорк: Вили. 115–175.
- 22. Mühl H, Kochem A-J, Disqué C, Sakka SG (2010) Активность и загрязнение ДНК коммерческих реагентов полимеразной цепной реакции для универсального обнаружения бактериальных патогенов в крови с помощью полимеразной цепной реакции 16 S рДНК в режиме реального времени. Диагностика Микр Инфек Дис 66: 41–49.
- 23. Раскин Л., Стромли Дж., Риттманн Б., Шталь Д. (1994) Зонды для групповой гибридизации 16 S рРНК для описания природных сообществ метаногенов. Appl Environ Microbiol 60: 1232–1240.
- 24. Шталь Д.А., Аманн Р. (1991) Разработка и применение зондов нуклеиновых кислот. В: Stackebrandt E, Goodfellow M, редакторы. Методы нуклеиновых кислот в бактериальной систематике. Нью-Йорк: Джон Уайли и сыновья. 205–248.
- 25. Хубер Х., Хон М.Дж., Рэйчел Р., Фукс Р., Виммер В.К. и др. (2002) Новый тип архей, представленный наноразмерным гипертермофильным симбионтом. Nature 417: 63–67.
- 26. Baker GC, Smith JJ, Cowan DA (2003) Обзор и повторный анализ доменно-специфичных праймеров 16 S.J Microbiol Methods 55: 541–555.
- 27. Gantner S, Andersson AF, Alonso-Sáez L, Bertilsson S (2011) Новые праймеры для анализа сообществ архей на основе 16 S рРНК в образцах окружающей среды. J Microbiol Methods 84: 12–18.
- 28. Pruesse E, Peplies J, Gloeckner FO (2012) SINA: точное высокопроизводительное выравнивание множественных последовательностей генов рибосомной РНК. Биоинформатика 28: 1823–1829.
- 29. Burggraf S, Stetter KO, Rouviere P, Woese CR (1991) Methanopyrus kandleri : архейный метаноген, не связанный со всеми другими известными метаногенами.Syst Appl Microbiol 14: 346–351.
- 30. Делонг Е.Ф. (1992) Археи в прибрежных морских средах. Proc Natl Acad Sci 89: 5685–5689.
- 31. Casamayor EO, Massana R, Benlloch S, Overeas L, Diez B и др. (2002) Изменения в сообществах архей, бактерий и эукариалов в зависимости от градиента солености путем сравнения методов генетического дактилоскопирования на графиках масштабирования с использованием нескольких водоемов. Appl Environ Microbiol 4: 338–348.
- 32. Brunk CF, Eis N (1998) Количественная мера последовательностей генов малых субъединиц рРНК Королевства Корархей.Appl Environ Microbiol 64: 5064–5066.
- 33. Embley TM, Finlay BJ, Thomas RH, Dyal PL (1992) Использование последовательностей рРНК и флуоресцентных зондов для исследования филогенетических положений анаэробной инфузории Metopus palaeformis и ее археобактериального эндосимбионта. J Gen Microbiol 138: 1479–1487.
- 34. Burggraf S, Huber H, Stetter KO (1997) Реклассификация порядков и семейств кренархов в соответствии с данными о последовательностях 16 S рРНК.Int J Syst Bacteriol 47: 657–660.
- 35. Schloss PD, Westcott SL, Ryabin T, Hall JR, Hartmann M, et al. (2009) Представляем mothur: программное обеспечение с открытым исходным кодом, независимое от платформы, поддерживаемое сообществом, для описания и сравнения микробных сообществ. Appl Environ Microbiol 75: 7537–7541.
- 36. Макдональд Д., Прайс М.Н., Гудрич Дж., Навроцкий Е.П., ДеСантис Т.З. и др. (2012) Улучшенная таксономия Greengenes с явными рангами для экологического и эволюционного анализа бактерий и архей.ISME J 6: 610–618.
- 37. Людвиг К., Странк О, Вестрам Р., Рихтер Л., Мейер Х и др. (2004) ARB: программная среда для данных последовательности. Nuc Acids Res 32: 136–1371.
- 38. Pruesse E, Quast C, Knittel K, Fuchs BM, Ludwig W и др. (2007) SILVA: всеобъемлющий онлайн-ресурс для проверенных и согласованных данных о последовательностях рибосомных РНК, совместимых с ARB. Nucleic Acids Res 35: 7188-7196.
- 39. Tourna M, Freitag TE, Nicol GW, Prosser JI (2008) Рост, активность и температурные реакции архей и бактерий, окисляющих аммиак, в почвенных микромирах.Environ Microbiol 10: 1357–1364.
- 40. Альтшул С.Ф., Гиш В., Миллер В., Майерс Е. В., Липман Д. Д. (1990) Базовый инструмент поиска локального выравнивания. J Mol Biol 215: 403–410.
- 41. Moissl C, Rudolph C, Huber R (2002) Природные сообщества новых архей и бактерий с морфологией, напоминающей жемчужины: молекулярный анализ бактериальных партнеров. Appl Environ Microbiol 68: 933–937.
- 42. Klappenbach JA, Saxman PR, Cole JR, Schmidt TM (2001) RrnDB: база данных количества копий оперона рибосомной РНК.Nucleic Acids Res 29: 181–184.
- 43. Mussmann M, Brito I, Pitcher A, Sinninghe Damsté JS, Hatzenpichler R, et al. (2011) Таумархеоты, изобилующие нитрифицирующими илами нефтеперерабатывающих заводов, экспрессируют amoA , но не являются обязательными автотрофными окислителями аммиака. Proc Natl Acad Sci USA 108: 16771–16776.
- 44. Флорес Г.Е., Бейтс С.Т., Найтс Д., Лаубер К.Л., Стромбо Дж. И др. (2011) Микробная биогеография поверхностей общественных туалетов. PLoS One 6: e28132.
- 45.Саудер Л.А., Петерс Ф.Ф., Схоутен С.С., Нойфельд Д.Д. (2012) Ниша аммиачных архей, окисляющих аммиак, во вращающихся биологических контакторах муниципальных очистных сооружений. Environ Microbiol 14: 2589–600.
- 46. Könneke M, Bernhard AE, de la TorreJR, Walker CB, Waterbury JB и др. (2005) Изоляция автотрофной морской археи, окисляющей аммиак. Природа 437: 543–546.
- 47. Lehtovirta-Morley LE, Stoecker K, Vilcinskas A, Prosser JI, Nicol GW (2011) Выращивание облигатного ацидофильного окислителя аммиака из нитрифицирующей кислой почвы.Proc Natl Acad Sci USA 108: 15892–15897.
- 48. Нос К., Мизуно Т., Ямане Н., Кондо Т., Охтани Х. и др. (2005) Определение аммиака в газе, исходящем из кожи человека, и его корреляция с газом в крови. Anal Sci 21: 1471–1474.
Адаптация цвета кожи человека в различных популяциях | Hereditas
С тех пор, как современные люди покинули Африку ~ 100 000 лет назад, они распространились по континентам в самых разных средах обитания, от тропических зон до Арктики и от низменностей до высокогорья.Во время миграции селективное давление в местных средах (например, холодный климат, гипоксия и эндемические патогены) вместе со случайным дрейфом привели к популяционным генетическим вариантам, которые в дальнейшем повлияли на различные фенотипы, такие как толерантность к лактозе, рост, иммунитет. система и метаболическая эффективность.
Вариация цвета кожи — один из наиболее ярких примеров фенотипического разнообразия человека. В нем преобладает меланин — пигментация, расположенная в основании эпидермиса и продуцируемая меланоцитами.Меланин имеет две формы: феомеланин (желто-красноватый) и эумеланин (черно-коричневый). Первые в основном накапливаются у светлокожих людей, а вторые — у смуглых [1,2,3,4,5]. Кроме того, количество и размер частиц меланина различаются у разных людей и даже более важны, чем пропорции двух форм меланина при определении цвета кожи человека [5]. Другие факторы, связанные с кожей, например кератин, также влияют на изменение цвета кожи [6, 7].
В глобальных популяциях цвет кожи сильно коррелирует с географической широтой и, в основном, с распределением ультрафиолетового (УФ) излучения (рис. 1). У населения, расположенного ближе к экватору, как правило, темная кожа для защиты от ультрафиолета, поскольку чрезмерное воздействие ультрафиолета может снизить уровень фолиевой кислоты [8, 9] и вызвать рак кожи [10,11,12,13]. Более светлая кожа у популяций в более высоких широтах лежит в основе отбора для поддержания фотосинтеза витамина D, который является УФ-зависимым процессом [14, 15].
Фиг.1Корреляция между цветом кожи и широтой (из Barsh (2003) [5]). ( a ) Карта распределения цвета кожи человека. ( b ) График зависимости коэффициента отражения кожи от широты
Хотя УФ считается движущей силой эволюции цветов кожи человека, понимание точного генетического механизма отбора будет иметь решающее значение для реконструкции истории эволюции человека и выяснения микроэволюции адаптивных черт. Описание полной картины региональной адаптации цвета кожи у людей было бы сложной задачей, поскольку она включает не только гены, идентифицированные как находящиеся в процессе отбора, но также и степень, в которой эти гены могут объяснить фенотипические вариации, взаимодействия и совместные эффекты генов, а также как они реагируют на внешнюю среду.В этой статье мы рассмотрели несколько случаев адаптации цвета кожи в различных популяциях современных людей и архаичных гомининов. Эти случаи показывают сходство и различие механизмов адаптации цвета кожи в разных популяциях и дают некоторое представление об истории эволюции человека.
Адаптация цвета кожи у современных евразийцев
У европейцев SLC24A5 и SLC45A2 [16,17,18,19] — два золотых гена, связанных с эволюцией светлого цвета кожи. SLC24A5 кодирует белок NCKX5, который является членом семейства трансмембранных белков и регулирует концентрацию кальция в меланосоме [16]. Было подтверждено, что этот ген влияет на пигментацию у рыбок данио и мышей [16, 20]. В частности, производный аллель rs1426654 в SLC24A5 оказался почти фиксированным у европейцев, но почти отсутствующим в популяциях без какого-либо европейского происхождения (рис. 2) [21]. Гаплотип размером 78 т.п.н. около SLC24A5 , который находится в неравновесном сцеплении с rs1426654, также был идентифицирован для накопления у европейцев [22].Сходный паттерн можно наблюдать на rs168
в SLC45A2 [23], который, как сообщается, связан с пигментацией у нескольких видов, например, мышей, рыб, птиц и лошадей [24,25,26]. Другие варианты этого гена, включая rs26722, rs2287949 и rs40132, также оказались ассоциированными с окраской у европейцев [23, 27, 28]. Другой важный ген, связанный с пигментацией, идентифицированный в Европе, — это MC1R [29,30,31]. Этот ген экспрессируется в меланоцитах и играет ключевую роль в контроле перехода с феомеланина на эумеланин [31].Пигментные фенотипы, связанные с MC1R , были изучены на широком круге животных [32,33,34]. Многие варианты были идентифицированы в MC1R , такие как rs1805007, rs1805008 и rs3212357 [35, 36], несмотря на его небольшой размер (951 п.н.). Другие важные европейские специфические локусы включают rs1393350 в TYR , rs2733831 в TYRP1, и rs18 в OCA2 [17, 28, 37,38,39]. Полученные частоты аллелей в этих локусах высоки у европейцев, но низкие у африканцев и выходцев из Восточной Азии, что может быть четким сигналом положительного отбора у европейцев, как показывает статистический анализ [40]. Рис. 2Эволюционная модель пигментации человека в трех континентальных популяциях. Укоренившееся дерево показывает генетическую филогению человеческих популяций из Африки, Северной Европы и Восточной Азии, а цвета ветвей примерно указывают на общий уровень пигментации кожи этих популяций (адаптировано из McEvoy et al. (2006) [39]). Генетические локусы, которые, как сообщается, находились под положительным отбором у общего предка современных евразийцев, представлены rs1881227 в KITLG , а те, которые независимо развились у европейцев и жителей Восточной Азии, что указывает на возможную конвергентную эволюцию, представлены rs122 в OCA2 и rs885479 в MC1R соответственно.Карты частоты аллелей были составлены с использованием R (версия 3.2.1, https://www.r-project.org) на основе этих локусов в 53 глобальных популяциях, предоставленных Группой по разнообразию генома человека CEPH (HGDP, http : //www.hagsc.org/hgdp/index.html). Синий и красный цвета обозначают предковые и производные аллели соответственно
.Гены, участвующие в адаптации цвета кожи у жителей Восточной Азии, не так хорошо изучены по сравнению с длинным списком адаптивных генов, идентифицированных у европейцев.Известные примеры включают OCA2 и MC1R . Каждый содержит несколько несинонимичных мутаций, например rs1800414 и rs74653330 в OCA2, и rs885479 в MC1R [40,41,42,43], которые демонстрируют высокие частоты производных аллелей у жителей Восточной Азии, но низкие частоты производных аллелей у европейцев и африканцев (рис. 2). Белок OCA2 считается зрелым мембранным белком меланосом [44], который может играть роль в транспортировке белка в меланосомы [45]. Восточноазиатский вариант rs1800414 был впервые описан в исследовании секвенирования экзома, целью которого было выявить варианты, связанные с альбинизмом [46].Производный аллель rs1800414, как полагали, вносил вклад в осветление кожи в исследовании ассоциации китайцев хань, которое измеряло цвет кожи людей с использованием индекса меланина [47]. Другой несинонимичный вариант в OCA2 , rs74653330, также был подтвержден как связанный с пигментацией в ассоциативном исследовании японцев [48]. Дополнительные примеры восточноазиатских специфичных аллелей, связанных с пигментацией, включают rs10809814 в TYRP1 и rs1407995 в DCT [40, 49], оба из которых демонстрируют различие между азиатами и неазиатами [47], а также сильные сигналы положительного отбора. у азиатов [43, 49].
Несмотря на различные гены и варианты при соответствующих местных адаптациях у европейцев и жителей Восточной Азии, некоторые гены имеют аллели, достигающие высоких частот в обеих континентальных группах. Например, KITLG демонстрирует избирательный поиск у неафриканцев [50,51,52]. Этот ген широко экспрессируется во многих тканях, включая кожу, и участвует в морфогенезе органов и пролиферации клеток. Kit-лиганд, кодируемый KITLG , известен как стальной фактор и играет решающую роль в нормальном развитии и поддержании линии меланоцитов во взрослой коже [53]; это было доказано на людях, рыбах и мышах [54,55,56].Влияние этого гена на пигментацию также было подтверждено в серии ассоциативных исследований [57,58,59,60]. Одним из ключевых вариантов является rs642742, который расположен на расстоянии 326 т.п.н. выше сайта начала транскрипции KITLG . При этом варианте частота предковых аллелей у африканцев превышает 90%, что сравнимо с полученной частотой аллелей у европейцев и выходцев из Восточной Азии (рис. 2). Подобные закономерности наблюдались и в других генах, например, ASIP и BNC2 [39].
Две модели эволюционной архитектуры пигментации человека были предложены на основе приведенных выше результатов и других связанных исследований (рис. 2). Одна из них — модель конвергентной эволюции [17, 40, 43, 49], предполагающая, что депигментация в некоторой степени развивалась независимо у европейцев и жителей Восточной Азии, поскольку для объяснения светлой кожи и положительного отбора у европейцев были предложены разные гены и варианты. эти две континентальные группы. По оценкам недавнего исследования, время выборочного обследования для специфичных для Европы вариантов пигментации было около 11 000–19 000 лет назад, после расхождения европейцев и азиатов [61].Альтернативная модель подходит для общих выборочных зачисток европейцев и выходцев из Восточной Азии, которые, возможно, могут произойти у прото-евразийцев. Начало этого движения было оценено приблизительно 30 000 лет назад, сразу после миграции «из Африки», но раньше, чем специфическая для Европы эволюция пигментации [61]. Сосуществование этих двух моделей предполагает сложную эволюционную историю цвета кожи у современных людей.
Еще одним ключом к разгадке сложной генетической основы эволюции цвета кожи является аллельная гетерогенность, наблюдаемая в одном гене, таком как OCA2 и MC1R .В каждом из этих генов одни аллели специфичны для европейцев, тогда как другие специфичны для азиатов, хотя доказано, что все они связаны с депигментацией. Кроме того, OCA2 предоставляет свидетельства независимых зачисток, а также конвергентной эволюции у европейцев и азиатов. Поскольку результаты были получены в результате исследований с использованием разных выборок, данных и методов, могут существовать некоторые смешивающие факторы, приводящие к этим различным наблюдениям. Однако, что более важно, цвет кожи — это сложная характеристика, которую нельзя просто объяснить одним геном или вариантом; скорее всего, это будет связано с огромной сетью генов и фенотипов.Например, ASIP , ген адаптивной пигментации в популяциях европейского происхождения [62, 63], кодирует сигнальный белок агути, который блокирует MC1R в синтезе эумеланина в ответ на УФ-индуцированное повреждение ДНК [40]. При производстве меланина TYR действует как катализатор ключевой начальной стадии, и его стабильность поддерживается TYRP1 и DCT .
Кроме того, сканирование для отбора по пигментации кожи указывает на два разных поведения отбора, действующих на мутации de novo и вариации стояния, соответственно.Некоторые варианты, представленные rs1805007 и rs1805008 в MC1R (у европейцев) и rs1800414 в OCA2 (у азиатов), показывают производные аллели только в популяциях с положительным отбором в этих локусах, из чего мы можем предположить, что они являются новыми мутациями. которые появились после того, как современные люди поселились в Европе или Азии. Напротив, некоторые варианты, такие как rs3212357 в MC1R (при положительном отборе у европейцев), имеют низкие частоты у африканцев. Независимо от возможных мутаций и генетического дрейфа в африканских популяциях, более вероятно, что производный аллель в этом локусе представлен в течение некоторого времени, прежде чем они стали популярными.Подобные случаи были обнаружены в высотной адаптации тибетцев и адаптации иммунитета в некоторых современных человеческих популяциях и даже в эволюции фенотипов пигментации у нечеловеческих видов [56, 64].
Адаптация цвета кожи в смешанных популяциях
Смешанные популяции, гибридное потомство двух ранее изолированных популяций, может дать важную информацию для понимания генетики географической изменчивости по двум причинам. Во-первых, локусы, лежащие в основе фенотипических различий в предковых популяциях, также перекрывают высокоинформативные маркеры происхождения, что делает смешанные популяции особенно полезными для отслеживания истории популяций.Во-вторых, смешанные популяции обычно имеют широкий диапазон вариаций в отношении некоторых конкретных фенотипов, что может повысить эффективность определения местоположения генов, связанных со сложными признаками / заболеваниями, после контроля потенциальной стратификации популяции.
Несмотря на эти преимущества, смешанные популяции редко учитывались в исследованиях изменения пигментации человека. Текущие исследования, посвященные изучению генов пигментации в смешанных популяциях, в основном касались лиц африканского и европейского происхождения, таких как афроамериканцы, европейцы-африканцы и латиноамериканцы, поскольку их предки существенно различаются по цвету кожи.Наследственные генетические составы этих трех популяций различаются. Афроамериканцы получили наибольший генетический вклад (~ 80%) от африканского происхождения [65], латиноамериканские метисы имеют наименьшую долю африканского происхождения (~ 10%) [66, 67], в то время как у европейских африканцев генетические компоненты унаследованные от европейцев (~ 42%) и африканцев (~ 58%) сопоставимы [68]. Только среди латиноамериканцев существует значительная часть индейского происхождения (~ 45%) [66, 67]. Более того, на индивидуальном уровне доля каждого предка в каждой смешанной популяции сильно различается.Например, доля европейского происхождения среди афроамериканцев колеблется от 2% до 98% [65]. Большая разница в цвете кожи у смешанных особей может быть результатом их очень разнообразного генетического состава, поскольку наблюдалась существенная корреляция между пропорциями предков и цветом кожи [68,69,70].
Множественные хорошо известные гены-кандидаты на пигментацию у европейцев также были идентифицированы с помощью картирования примесей (рис. 3) или ассоциативных исследований в смешанных популяциях.Например, TYR , несущий несинонимичную замену rs1042602 (S192Y), был идентифицирован у афроамериканцев [69] и европейских африканцев из Кабо-Верде [68]. Варианты ASIP, , такие как rs6058017, который, как было обнаружено, встречается с разной частотой в глобальных популяциях [63], также были связаны с темными волосами и карими глазами у американцев европейского происхождения [71], афроамериканцев [62]. и бразильцы [72]. Более того, KITLG продемонстрировал сильные сигналы избирательного захвата у афроамериканцев [51] со значительным предпочтением гомозиготам по афро-специфическому аллелю (предковому аллелю) по rs642742 у людей с высоким индексом меланина (темная кожа) [69].Подобные случаи включают rs1426654 в SLC24A5 у европейских африканцев [68] и латиноамериканцев [72] и rs35395 в SLC45A2 у европейских африканцев [68].
Рис. 3Структура картирования примесей для обнаружения положительного отбора. Средние лица африканцев, европейцев и африканцев были загружены с http://www.mediadump.com/hosted-id167-average-faces-from-around-the-world.html#.WLkMU-kfU1A
Однако в некоторых исследованиях получены противоречивые результаты.Корреляция между коренным американским происхождением и пигментацией кожи, о которой сообщалось у латиноамериканского населения [73], не наблюдалась в группе пуэрториканских женщин [70]. Один из ключевых локусов однонуклеотидного полиморфизма (SNP) в OCA2 , rs1800404, показал значительное влияние на пигментацию кожи при анализе афроамериканцев и объединенной популяции афроамериканцев и афро-карибских стран, но отсутствовал в независимом анализе Афро-карибские образцы [69]. Возможно, что в разных популяциях существуют разные генетические механизмы изменения цвета кожи, но следует проявлять осторожность в отношении подробной информации в данных, такой как размер выборки и предковые популяции, выбранные для анализа смешанных популяций, что может привести к смещенным результатам [ 67, 69].
Идентификация генетических детерминант естественной изменчивости пигментации кожи проводилась также в других смешанных популяциях. Одним из успешных примеров является полногеномное исследование ассоциации населения южноазиатского происхождения [74], в котором полиморфизмы в SLC24A5 , TYR и SLC45A2 показали значительную связь с содержанием меланина в коже. Аллели светлой кожи в Южной Азии могли быть унаследованы от их европейских предков [75], которые первоначально прибыли в этот регион около 3500–4000 лет назад вместе с индоевропейской языковой экспансией [76], за которой последовала недавняя колонизация в последние несколько лет. веков.Кроме того, Центральная Азия и Юго-Восточная Азия являются домом для различных смешанных популяций, которые также имеют большой потенциал в изучении адаптации к цвету кожи. Смешанный популяционный анализ может значительно обогатить наше понимание вариаций цвета кожи в современных популяциях людей.
Адаптация цвета кожи в популяциях аборигенов
Популяции аборигенов в различных регионах мира имеют множество последствий для истории эволюции человека. Они считались первыми поселенцами в соответствующих областях.Несмотря на то, что некоторые аборигены в некоторой степени ассимилировались окружающими их земледельцами, они сохранили свои традиционные средства к существованию в качестве охотников-собирателей, а также свои изначальные физические черты — темную кожу, низкий рост и вьющиеся волосы.
Популяции охотников-собирателей с темной кожей, невысоким ростом и вьющимися волосами привлекли большое внимание (рис. 4а). Генетический механизм, лежащий в основе общих фенотипов среди этих географически удаленных популяций (вместе называемых негритосами или пигмеями) из Центральной Африки, Андаманских островов, Юго-Восточной Азии и Океании, все еще остается спорным; например, были ли они общим происхождением от донеолитического субстрата человечества или следствием конвергентной эволюции [77, 78].На сегодняшний день большинство генетических исследований по этому вопросу сосредоточено на росте [78,79,80,81]. Одно исследование предоставило ключи к разгадке конвергентной эволюции с точки зрения адаптации пигментации кожи, проанализировав разнообразие MC1R у меланезийцев [82]. Это исследование показало, что предковые гаплотипы MC1R не очень консервативны между меланезийцами северных островов и африканцами, хотя обе популяции живут в области с высоким УФ-излучением, что контрастирует с предыдущими выводами, основанными на очень ограниченных выборках [30, 83].Кроме того, несинонимичный полиморфизм, rs2228479, показывает обогащенные производные аллели, особенно у жителей Восточной Азии, но не имеет существенной связи с пигментацией кожи или волос у меланезийцев. Фактически, популяция меланезийцев демонстрирует поразительную вариабельность пигментации кожи [84], и, как всегда, некоторые варианты были идентифицированы как регионально-специфичные, что может частично объяснить эту фенотипическую вариацию. Ярким примером является несинонимичный вариант, rs387 1, в TYRP1 [85].Он ограничен Соломоновыми островами и частями архипелага Бисмарка и может способствовать появлению «светлых волос» в этом регионе [85, 86]. Эти результаты подчеркивают сложную генетическую архитектуру фенотипов пигментации, а также подчеркивают роль, которую популяционная история (например, сложная популяционная история юго-западной части Тихого океана [87,88,89]) может играть роль во влиянии на фенотипическое разнообразие. Исследования пигментации кожи у других современных аборигенных популяций (помимо меланезийцев) немногочисленны, за исключением одного, изучающего популяцию сеноев (коренное население) с Малайского полуострова, которое представляет собой смесь негрито (темнокожих) и южных монголоидов из Индо. -Китай (с желто-коричневой кожей) и имеет широкий спектр цветов кожи [90].Авторы этого исследования обнаружили, что, несмотря на низкую частоту производных аллелей, мутация A111T (rs1426654) в SLC24A5 в значительной степени связана со светлой кожей у сеноев, которая, как предполагалось, возникла в результате смешения монголоидов и жителей Южной Азии.
Рис. 4Цвет кожи аборигенов Экваториальной зоны и Арктики. ( a ) Сравнение цвета кожи между Bateq (подгруппа Negrito) и малайцем из полуостровной Малайзии. ( b ) Сравнение цвета кожи инуитов и шведов из схожих широт.Портреты малайцев и шведов предоставлены проектом Джошуа (http://joshuaproject.net), портрет Батека взят с сайта http://www.businessinsider.my/, а портрет инуитов с сайта http: // www. arcticphoto.co.uk/
Другой интересный вопрос, связанный с адаптацией цвета кожи человека, исходит от жителей Арктики. У инуитов в Дальнем Северо-Востоке Азии и в Американской Субарктике кожа желтовато-коричневая, несмотря на крайнюю северную широту, на которой они живут, в отличие от других популяций, живущих на той же широте, таких как шведы и финны (рис.4б). Это делает инуитов исключением из коррелированного с широтой распределения цвета кожи. Одна из возможных причин заключается в том, что темная кожа могла защитить инуитов от сильного воздействия ультрафиолета из-за долгого светового дня зимой и высокого уровня отражения ультрафиолета от снега. В то время как темная кожа является недостатком для производства витамина D, большое количество витаминов, включая витамин D, может быть компенсировано их диетой [91, 92]. Другой причиной может быть эффект основателя древних восточноазиатских предков инуитов, которые населяли арктический регион почти 5000 лет назад и имели более высокое производство меланина, чем европейские предки.Однако было проведено очень мало генетических исследований для определения генетической основы темной кожи в арктических популяциях.
Адаптация цвета кожи у древних гомининов
Темная кожа современных людей возникла около 1,2 миллиона лет назад в результате потери волос на теле после расхождения с обезьянами, предположительно для защиты от повреждений, вызванных ультрафиолетом [13, 93, 94,95,96]. Тогда, когда же у современных евразийцев началась депигментация? Обобщенные выше исследования адаптации цвета кожи основаны на современных популяционных генетических данных, которые могут страдать от ограниченного временного разрешения, вызванного демографической историей популяции, и нечувствительностью к отбору, действующему на вариации стояния [97].Появление древних анализов ДНК позволяет непосредственно наблюдать за процессами эволюции и, таким образом, облегчить нам понимание этого ключевого вопроса.
Исследование геномов анатолийских неолитических фермеров в Западной Евразии (6500–300 гг. До н.э.), которые, вероятно, являются исходным населением первых европейских фермеров, предполагает, что светлый цвет кожи возник по крайней мере с 6500–4000 лет назад. [98]. Несколько популярных генов, идентифицированных у современных евразийцев, например, SLC45A2 , GRM5 и HERC2 / OCA2 , показали сильный сигнал отбора в этих древних образцах.Этот вывод подтверждается другим исследованием, основанным на образцах энеолита (6500–5000 лет до н.э.) и бронзового века (5000–4000 лет назад), представляющих ранних европейских фермеров или поздних охотников-собирателей в Центральной Европе [99]. Одной из возможных причин депигментации кожи в доисторической Евразии является сельское хозяйство, которое привело к переходу с богатой витамином D диеты охотников-собирателей на диету земледельцев с низким содержанием витамина D, а также возросшая опасность дефицита фолиевой кислоты в более высоких широтах [14 , 100].Более того, давление отбора продолжало действовать долгое время после того, как оно инициировало адаптацию цвета кожи, поскольку некоторые аллели предковой пигментации были идентифицированы у европейцев мезолита (7000 лет назад), а некоторые адаптивные аллели отбора у древних евразийцев все еще развиваются. у современного человека [98, 99, 101].
Недавние исследования архаичных гомининов (например, неандертальцев, вымершая группа гоминидов, жившая в Евразии ~ 400–28000 лет назад [102]) еще больше улучшили наше понимание эволюции цвета кожи у современных людей.Неандертальцы встретили современных людей на Ближнем Востоке ~ 60 000–50 000 лет назад и внесли свой вклад примерно в 1–4% геномов современного человека [103,104,105]. Некоторые гены, связанные с пигментацией, идентифицированы в интрогрессированных гаплотипах неандертальцев современных евразийцев, таких как POU2F3 , BNC2 и MC1R [106, 107]. В частности, сообщалось, что интрогрессивные аллели приводят к светлому цвету кожи, что указывает на стратегию «адаптивной интрогрессии» адаптации цвета кожи человека.Другие интрогрессивные гены, связанные с фенотипами кожи, включают гены HYAL , которые связаны с клеточными ответами на УФ и находятся под сильным положительным отбором у жителей Восточной Азии [108], а также гены, участвующие в образовании кератиновых волокон [109]. Хотя эти гены не являются прямыми детерминантами пигментации кожи, они, как и гены, связанные с пигментацией, возможно, помогли современным людям адаптироваться к неафриканской среде.
Делая выводы об адаптивной интрогрессии, мы фактически утверждаем, что неандертальцы могли иметь светлый цвет лица.Этот вывод основан только на некоторых связанных с пигментацией генах или аллелях, идентифицированных в существующих современных человеческих популяциях, поскольку видимые фенотипы неандертальцев и других вымерших видов недоступны. Однако при использовании некоторых других априорных генов в качестве потенциальных ключей можно получить другие результаты. Например, производное состояние MC1R , которое отвечает за бледность кожи, присутствует у неандертальцев из Италии и Испании, но отсутствует у хорватских неандертальцев и Денисовой [110], что свидетельствует об изменении цвета кожи у архаичных гомининов.Кроме того, светлая кожа у неандертальцев и современных евразийцев также могла быть результатом конвергентной эволюции, а не адаптивной интрогрессии [111].
Гипотеза об адаптивной интрогрессии, похоже, возникла раньше, когда современный человек побледнел — задолго до эпохи позднего мезолита, когда неандертальцы вымерли около 28000 лет назад. Однако мы должны пересмотреть, действительно ли гены, влияющие на цвет кожи архаичных гомининов, определяли цвет кожи у современных людей. Даже если это так, также возможно, что современный человек сохранил эти интрогрессивные варианты до тех пор, пока они не проявили некоторые фенотипические эффекты при определенных сильных селективных давлениях.Таким образом, для решения этой проблемы в будущем необходимы дополнительные ресурсы данных и анализы.
Коэффициент отбора и величина эффекта
Как одно из наиболее очевидных изменений в окружающей среде после миграции современного человека из Африки в более высокие широты, УФ оказывает значительное селективное давление на пигментацию кожи человека, что может быть отражено коэффициентами отбора гены, связанные с пигментацией. Оценка коэффициентов отбора во многом зависит от рассматриваемых генов и методологий.Beleza et al. оценили коэффициент отбора в нескольких локусах, представляющих SLC24A5 , SLC45A2 , TYRP1, и KITLG [61]. Например, оценки составляют 0,05 / 0,04 для SLC45A2 и 0,16 / 0,08 для SLC24A5 при доминантной / аддитивной модели наследования у европейцев. Между тем, López et al. сообщили, что коэффициент отбора варианта в SLC45A2 составляет 0,01–0,02 в южноевропейских популяциях [112].Эти оценки сопоставимы с коэффициентами отбора, выведенными непосредственно из данных с последовательной выборкой в HERC2 , SLC45A2 и TYR , в диапазоне 0,02–0,1 [99]. Коэффициенты отбора, оцененные для генов пигментации, лучше всего понять в контексте оценок для других недавно выбранных локусов. Преимущество отбора, как предполагается, составляет 0,01–0,08 для LCT , гена, сильно связанного с персистенцией лактазы в популяциях европейского происхождения [113, 114], 0.019–0,048 для G6PD , гена, обеспечивающего устойчивость к малярии в африканских популяциях [115], 0,03–0,19 для EDAR , связанного с увеличением толщины волос на скальпе и изменением морфологии зубов у китайцев хань [116], и 0,0004–0,0023 для областей гена EGLN1 и EPAS1 , вносящих вклад в высотную адаптацию тибетцев [117]. Коэффициенты отбора для генов пигментации являются одними из наиболее тщательно отобранных генов в геноме человека, что указывает на серьезное давление отбора, вызванное УФ-излучением или некоторыми другими изменениями окружающей среды в неафриканских регионах.
Хотя было обнаружено, что большое количество генов вносит вклад в изменение цвета кожи, насколько они могут объяснить изменение цвета кожи у современных людей? Есть ли ген или вариант, который оказывает доминирующее влияние на цвет кожи? Некоторые гены могут играть важную роль в определении цвета кожи в определенных популяциях. Например, вариант со светлой кожей на rs1426654 в SLC24A5 может объяснить 22–32% дисперсии индекса меланина в Южной Азии [75] и 25–38% в популяциях афроамериканцев и афро-карибских народов [118].Кроме того, производный аллель rs642742 в KITLG может объяснить осветление кожи человека на 6-7 единиц меланина, что составляет почти 1/5 от общей разницы в отражательной способности кожи между западноафриканцами и европейцами (30 единиц меланина) [56]. Однако существует относительно больше генов и вариантов с меньшими эффектами. Один из ключевых вариантов в OCA2 , rs1800414, может объяснять около 4% вариаций пигментации в популяциях Восточной Азии [47]. В странах Южной Азии rs168
в SLC45A2 и rs1042602 в TYR составляют 3.Изменение цвета кожи на 6% и 2,5%, соответственно, намного меньше, чем размер эффекта rs1426654 в SLC24A5 [74]. Тип наследования пигментации кожи следует аддитивной модели или, по крайней мере, неполной аддитивной модели [16, 17, 47, 56, 75].Клетки кожи человека трансформируются непосредственно в двигательные нейроны — Медицинская школа Вашингтонского университета в Сент-Луисе
Посетите новостной центрНовая техника может помочь в лечении заболеваний, ведущих к параличу
Дэниел АбернатиУченые открыли новый способ превращения клеток кожи человека непосредственно в двигательные нейроны (см. Выше).Методика, разработанная в Медицинской школе Вашингтонского университета в Сент-Луисе, может помочь исследователям лучше понять болезни двигательных нейронов, такие как боковой амиотрофический склероз. Моторные нейроны человека трудно изучать, так как их нельзя получить от живых пациентов. Изображенные двигательные нейроны были преобразованы из клеток кожи, взятых у здоровой 42-летней женщины.
Ученые, работающие над разработкой новых методов лечения нейродегенеративных заболеваний, оказались в тупике из-за невозможности выращивать человеческие двигательные нейроны в лаборатории.Моторные нейроны вызывают мышечные сокращения, и их повреждение лежит в основе разрушительных заболеваний, таких как боковой амиотрофический склероз и спинальная мышечная атрофия, которые в конечном итоге приводят к параличу и ранней смерти.
В новом исследовании ученые из Медицинской школы Вашингтонского университета в Сент-Луисе превратили клетки кожи здоровых взрослых людей непосредственно в двигательные нейроны, минуя состояние стволовых клеток.
Методика позволяет исследовать мотонейроны центральной нервной системы человека в лабораторных условиях.В отличие от обычно изучаемых двигательных нейронов мышей, двигательные нейроны человека, растущие в лаборатории, будут новым инструментом, поскольку исследователи не могут брать образцы этих нейронов у живых людей, но могут легко брать образцы кожи.
Исследование опубликовано 7 сентября в журнале Cell Stem Cell.
Избегание фазы стволовых клеток устраняет этические проблемы, возникающие при производстве так называемых плюрипотентных стволовых клеток, которые похожи на эмбриональные стволовые клетки по своей способности превращаться во все типы взрослых клеток.И что важно, предотвращение состояния стволовых клеток позволяет образовавшимся мотонейронам сохранять возраст исходных клеток кожи и, следовательно, возраст пациента. Сохранение хронологического возраста этих клеток жизненно важно при изучении нейродегенеративных заболеваний, которые развиваются у людей в разном возрасте и ухудшаются в течение десятилетий.
«В этом исследовании мы использовали клетки кожи только здоровых взрослых людей в возрасте от 20 до 60 лет», — сказал старший автор Эндрю С. Ю, доктор философии, доцент кафедры биологии развития.«Наше исследование показало, как небольшие молекулы РНК могут работать с другими клеточными сигналами, называемыми факторами транскрипции, для генерации определенных типов нейронов, в данном случае моторных нейронов. В будущем мы хотели бы изучить клетки кожи пациентов с нарушениями двигательных нейронов. Наш процесс преобразования должен моделировать аспекты заболевания с поздним началом, используя нейроны, полученные от пациентов с этим заболеванием ».
«Возвращение через фазу плюрипотентных стволовых клеток немного похоже на снос дома и постройку нового с нуля», — сказал Ю.«То, что мы делаем, больше похоже на ремонт. Мы меняем интерьер, но оставляем исходную структуру, которая сохраняет характеристики стареющих взрослых нейронов, которые мы хотим изучить ».
Способность ученых преобразовывать клетки кожи человека в клетки других типов, такие как нейроны, может улучшить понимание болезней и привести к поиску новых способов исцеления поврежденных тканей и органов — области, называемой регенеративной медициной.
Daniel AbernathyКлетки кожи человека (см. Выше), взятые у здорового взрослого человека и затем преобразованные в нейроны различных типов, могут стать ценным инструментом исследования.Подобные образцы кожи пациентов с нейродегенеративными заболеваниями могут позволить ученым изучить болезнь на ее естественном типе клеток.
Чтобы преобразовать клетки кожи в двигательные нейроны, исследователи подвергли клетки кожи молекулярным сигналам, которые обычно присутствуют на высоких уровнях в головном мозге. Прошлая работа Ю и его коллег — тогда еще из Стэнфордского университета — показала, что воздействие двух коротких фрагментов РНК превращало клетки кожи человека в нейроны. Эти две микроРНК, называемые miR-9 и miR-124, участвуют в переупаковке генетических инструкций клетки.
В новом исследовании исследователи подробно охарактеризовали этот процесс переупаковки, подробно описав, как клетки кожи, перепрограммированные в обычные нейроны, затем могут быть направлены в определенные типы нейронов. Они обнаружили, что гены, участвующие в этом процессе, готовы к экспрессии, но остаются неактивными до тех пор, пока не будет обеспечена правильная комбинация молекул. После долгих экспериментов с множеством комбинаций исследователи обнаружили, что добавление еще двух сигналов — факторов транскрипции, называемых ISL1 и LHX3, — примерно за 30 дней превратило клетки кожи в мотонейроны спинного мозга.
Комбинация сигналов — микроРНК miR-9 и miR-124 плюс факторы транскрипции ISL1 и LHX3 — говорит клетке свернуть генетические инструкции для создания кожи и развернуть инструкции для создания моторных нейронов, согласно Ю и соавторам исследования. первые авторы, Дэниел Г. Абернати и Мэтью Дж. Маккой, докторанты в лаборатории Ю; и У Гён Ким, доктор философии, научный сотрудник постдокторантуры.
Другое прошлое исследование, проведенное командой Ю, показало, что воздействие одних и тех же двух микроРНК, miR-9 и miR-124, а также разного сочетания факторов транскрипции может превратить клетки кожи в нейрон другого типа.В этом случае клетки кожи превратились в нейроны полосатого тела со средними шипами, которые поражены болезнью Хантингтона — наследственным, в конечном итоге фатальным генетическим заболеванием, которое вызывает непроизвольные движения мышц и когнитивные нарушения, начинающиеся в среднем зрелом возрасте.
В новом исследовании исследователи заявили, что преобразованные мотонейроны выгодно отличаются от нормальных моторных нейронов мыши с точки зрения включаемых и выключаемых генов и того, как они функционируют. Но ученые не могут быть уверены, что эти клетки идеально подходят для естественных мотонейронов человека, поскольку сложно получить образцы культивируемых мотонейронов у взрослых людей.Дальнейшая работа по изучению образцов нейронов, полученных от пациентов после смерти, необходима, чтобы определить, насколько точно эти клетки имитируют естественные двигательные нейроны человека.
Эта работа поддержана Премией новатора Национального института здравоохранения (NIH), грант номер DP2NS083372-01; Президентская премия за раннюю карьеру для ученых и инженеров; Программа исследования травм / заболеваний спинного мозга в штате Миссури; Фонд лечения болезни Альцгеймера; Стипендия факультета Эндрю Б. и Вирджинии К. Крейг; стипендия Филиппа и Симы Нидлман для аспирантов; a Рут Л.Институциональная стипендия Национальной исследовательской службы Киршштейна, номер гранта T32GM081739; и Детский институт открытий.
Абернати Д.Г., Ким В.К., Маккой MJ, Лейк AM, Оувенга Р., Ли С.В., Син Икс, Ли Д., Ли Х.Дж., Хекерот Р.О., Догерти Д.Д., Ван Т., Ю А.С. МикроРНК индуцируют пермиссивную среду хроматина, которая позволяет нейрональному подтипу репрограммировать фибробласты взрослого человека. Стволовая клетка. 7 сентября 2017 г.
Медицинский факультет Вашингтонского университета насчитывает 2100 штатных и добровольных врачей-преподавателей, которые также входят в медицинский персонал Barnes-Jewish и St.Детские больницы Луи. Медицинский факультет — одно из ведущих медицинских исследовательских, учебных и лечебных учреждений в стране, которое в настоящее время занимает седьмое место в стране по версии U.S. News & World Report. Медицинская школа связана с BJC HealthCare через свои связи с больницами Barnes-Jewish и St. Louis Children’s.
границ | Модель кожи человека ex vivo для изучения поверхностных грибковых инфекций
Введение
Кожные грибковые инфекции (SFI) поражают 25% населения мира (Pires et al., 2014; Чжань и Лю, 2016). Большинство этих инфекций являются поверхностными, что означает, что они поражают только эпидермис, самый внешний слой кожи (Chu et al., 2003). Другие SFI классифицируются как подкожные и глубокие, которые встречаются с гораздо меньшей частотой, но имеют значительную заболеваемость и смертность (Queiroz-Telles et al., 2017).
Дерматофиты вызывают поверхностные инфекции и в зависимости от среды обитания классифицируются как зоофильные, геофильные или антропофильные. Зоофильные виды могут инфицировать людей, но инфекция обычно вызывает сильное воспаление у людей, тогда как у животных она может протекать бесследно (Chu et al., 2003). Антропофильные виды, колонизирующие и адаптированные к людям, могут вызывать необычные случаи дерматофитии у животных, клинические проявления которых отличаются от клинических проявлений человека (Brilhante et al., 2006). Различия в клинической картине между разными видами представляют собой главное ограничение животных моделей для изучения SFI.
Ранее для изучения SFI использовалось несколько моделей, таких как кератино-соевая среда (Zaugg et al., 2009), слои рогового слоя (Peres et al., 2016), культура клеток монослоя кератиноцитов (Huang et al., 2015) и реконструированный эпидермис человека (Liang et al., 2016; Faway et al., 2017). Ограничения этих упомянутых моделей включают отсутствие иммунных клеток или кератиноцитов. Кроме того, кератиноциты обычно не погружаются в среду для культивирования тканей, а поверхность кожи обычно подвергается воздействию внешней среды и воздуха.
Ex vivo эксплантатов кожи, полученных в результате хирургических вмешательств, обычно восстановительных, были использованы для исследования восстановления кожного барьера (Danso et al., 2015), заживление ран (Xu et al., 2012), химическая токсичность (Nakamura et al., 1990), хронические воспалительные заболевания (Guilloteau et al., 2010), вакцинация ДНК (Ng et al., 2009) и грибковая инфекция. инфекции (Peres et al., 2016; Poyntner et al., 2016). В этих публикациях описаны различные методологии и применения модели эксплантата кожи. Используемые анализы включают гистологию, флуоресцентную микроскопию, иммунофлуоресценцию и иммуногистохимию для идентификации иммунных клеток и белков, электронную микроскопию, высокоэффективную тонкослойную хроматографию для измерения липидного состава и измерения экспрессии генов с помощью qRT-PCR и RNAseq.
Trichophyton rubrum , антропофильный вид грибов, является наиболее частой причиной поверхностных микозов (Havlickova et al., 2008). T. rubrum показывает спонтанное заживление на животных моделях, что отличается от того, как процесс заживления происходит у людей. Ответы хозяина против T. rubrum различаются в зависимости от модели, используемой для изучения взаимодействия хозяин-патоген. Например, экспрессия грибковых генов, связанных с вирулентностью ( acuD, citA, acuE, cys3, hexA ) и ростом грибов ( pacC, ssU, cdo ), выше, когда T.rubrum изучается в листах рогового слоя или росте на кератине, по сравнению с ex vivo человеческими эксплантами (Peres et al., 2016).
Необходимо подробное описание методологии создания модели кожи человека ex vivo . В опубликованных протоколах подробно описаны важные методологические различия, такие как разная толщина кожи, тип среды с добавками, размер образца, свежесть ткани, температура хранения и инкубации, а также частота смены среды (Nakamura et al., 1990; Сюй и др., 2012; Danso et al., 2015; Пойнтнер и др., 2016).
Мы определили наиболее часто используемые методы и анализы из предыдущих отчетов и установили стандартизированную методологию для изучения SFI. Мы сообщаем о нашем протоколе модели ex vivo кожи человека и результатах инфекции T. rubrum как лучшем примере первичной поверхностной инфекции кожи. Наша основная цель — предоставить модель SFI, которая точно отражает заболевание человека и может быть проанализирована на молекулярном уровне для исследования взаимодействий грибок: хозяин.Протокол включает подробное описание методологии подготовки образца кожи и установления инфекции, советы о том, как избежать заражения, и подробные сведения о том, как можно получить образцы высокого качества для последующих анализов.
Краткое описание процедуры следующее: конидий T. rubrum были выделены из культур агара, инкубированных в течение 7–10 дней при 30 ° C. Конидии определяли количественно и инокулят доводили до введения 1 × 10 6 клеток в 10 мкл PBS.
Хирургические кожные эксплантаты вскрыли для получения кусочков размером 1 см 2 . Поверхность каждого кусочка кожи осторожно пораняли иглой. Затем каждый кусочек кожи помещали в 6-луночный планшет и добавляли 1 мл среды с добавлением DMEM, всегда поддерживая межфазное пространство воздух-жидкость. Посевной материал грибка (10 мкл) добавляли на поверхность кожи, избегая контакта с дермой и окружающей средой. Отрицательный контроль (незараженная кожа) включали в каждый эксперимент. Чашки с культурами инкубировали при 37 ° C в 5% CO 2. Культуральную среду заменяли свежей средой каждые 24 часа, а отработанную среду хранили в пробирках на 2 мл при -80 ° C. Образцы тканей кожи были собраны в разные моменты времени для дальнейшего анализа.
Анализ полученных образцов включал сканирующую электронную микроскопию (SEM), гистологию, флуоресцентную микроскопию для оценки апоптоза и окрашивание калькофлуором в белый цвет для подтверждения грибковой инфекции и изучения морфологии грибка. Мы также описываем анализы для изучения SFI, такие как экстракция РНК и qRT-PCR для измерения экспрессии генов человека, а также экстракция белка из тканей и супернатантов для протеомного анализа с помощью жидкостной хроматографии-масс-спектрометрии (LC-MS / MS).
Материалы и оборудование
Культура грибов
— Картофельный агар с декстрозой (КПК), картофельный крахмал 4 г / л, декстроза / глюкоза 20 г / л. Термо Фишер. (Лафборо, Великобритания).
— Статический инкубатор. ООО «Лаборатория теплового оборудования LTE» (Олдхэм, Великобритания).
— Trichophyton rubrum штамм CBS 304.60. Вестердейкский институт биоразнообразия грибов.
— Камерный гемоцитометр Нейбауэра.
— Распределители ячеек L-образной формы (39 мм × 140 мм).Microspec Ltd. (Бромборо, Великобритания).
— Раствор фосфатного буфера Дульбекко, 500 мл. Сигма-Олдрич (Дорсет, Великобритания).
— Центрифуга Cellstar (пробирки 15 мл и 50 мл). Greiner Bio-One (Кремсмюнстер, Австрия).
Кожная процедура
— Кожная ткань человека без жирового слоя (жировой слой был удален хирургом в момент операции) и без растяжек после операций на брюшной полости или груди. Предоставлено Tissue solutions ® Ltd.(Глазго, Великобритания).
— Охладитель, сухой лед и / или охлаждающие пакеты для транспортировки.
— Стерильные щипцы, хирургические ножницы и скальпели.
— Стерильные хирургические лезвия, размер 15 из нержавеющей стали. Swann-Morton (Англия, Великобритания).
— Одноразовые стерильные иглы (19G, 2 ″, 1,1 × 50 мм). BD microlance 3. BD Biosciences (Дроэда, Ирландия).
— 6-луночный планшет Nunc Delta для культивирования клеток. Thermo Fisher Scientific.
— Среда Игла, модифицированная Дульбекко (DMEM).Thermo Fisher Scientific.
— Пенициллин 10000 ЕД / мл и стрептомицин 10 мг / мл, 100 мл. Thermo Fisher Scientific.
— Инактивированная нагреванием фетальная бычья сыворотка (HI-FBS). Thermo Fisher Scientific.
— Стерильные наконечники для пипеток TipOne 1000 мкл, 200 мкл, 10 мкл. Starlab (Милтон Кейнс, Великобритания).
— CO 2 лабораторный инкубатор.
— Нитриловые перчатки.
— Микроцентрифужные стерильные чистые пробирки, 2 мл и 1,5 мл.
— Чашки Петри из полистирола стерильные.Стерильно. Greiner Bio-one. (Кремсмюнстер, Австрия).
— Стереомикроскоп Carl Zeiss TM Stemi 2000-c. Carl Zeiss (Оберкохен, Германия).
— Дезинфицирующее средство высокого уровня Chemgene. СТАРЛАБ.
Гистология
— OCT Embedding Matrix Cellpath, Ltd. (Ньютаун, Великобритания).
— Одноразовые базовые формы (15 × 15 × 5 мм). Park Scientific (Нортгемптон, Великобритания).
— Сухой лед.
— Реактив лабораторный изопентан.Thermo Fisher Scientific.
— Слайды с покрытием из поли- L -лизина. Thermo Fisher Scientific.
— Криостат Leica CM1900. Leica Biosystems (Вецлар, Германия).
— Покровное стекло. 100 шт 22 × 50 мм. VWR International (Латтерворт, Великобритания).
— 16% параформальдегид, не содержащий метанола (мас. / Об.) 10 мл. Thermo Fisher Scientific.
— Модифицированное окрашивание серебром GMS, (HT-100), Sigma-Aldrich.
— Модифицированный раствор гематоксилина Харриса. Сигма-Олдрич.
— Светло-зеленый СФ желтоватый 10 гр. акрос органика. Thermo Fisher Scientific.
— Histoclear. Гистологический очищающий агент. Национальная диагностика (Ноттингем, Великобритания).
— Histomount. Гистологическая среда для закрепления. Национальная диагностика (Ноттингем, Великобритания).
— Zeiss TM Axio scan.Z1 Цифровой слайд-сканер. Карл Цейсс.
— Флуоресцентный отбеливатель 28 (Calcofluor white M2R). Сигма-Олдрич.
— Гидроксид калия (КОН).Сигма-Олдрич.
— Конфокальный микроскоп DeltaVision TM . GE Healthcare. (Бакингемшир Соединенное Королевство).
— Иодид пропидия> 94% ВЭЖХ. Сигма-Олдрич.
— DeadEnd TM Флуорометрическая система ТУНЕЛЬ. Промега (Саутгемптон, Великобритания).
— Монтажная среда Vectashield ® для флуоресценции с DAPI и без него, для окрашивания TUNEL и Calcofluor соответственно. Vector Laboratories (Питерборо, Великобритания).
— этанол 100%.
Сканирующая электронная микроскопия (СЭМ)
— 50% глутарового альдегида в H 2 O. Sigma-Aldrich.
— Тригидрат какодилата натрия. Сигма-Олдрич.
— Четырехокись осмия. Сигма-Олдрич.
— этанол 100%.
— Гексаметилдисилазан (HMDS). Сигма-Олдрич.
— Сканирующий электронный микроскоп Zeiss TM EVO MA10. Карл Цейсс.
qRT-PCR
— РНК позже ® Реагент для хранения тканей.Сигма-Олдрич.
— RNaseZap TM . Средство для удаления и очистки РНКазы. Сигма-Олдрич.
— раствор ТРИЗола ТМ . Thermo Fisher Scientific.
— Хлороформ.
— Изоамиловый спирт.
— Изопропанол.
— Азот жидкий.
— Стерильная ступка и пестик.
— FastPrep TM -24 5G MP Biomedicals.
— MP Biomedicals TM Пробирки Lysing Matrix D, керамические шарики 1,4 мм.MP Biomedicals.
— Диэтилпирокарбонат, 97% (DEPC). Сигма-Олдрич. Для очистки воды использовался 0,1% DEPC.
— Спектрофотометр NanoDrop TM 1000. Thermo Fisher Scientific.
— буфер 5X TBE (основание TRIS 54 г / л, борная кислота 27,5 г / л и 10 мМ EDTA сверхчистый). Рабочий раствор 1X TBE. Сделайте буфер, используя DEPC-воду.
— Без агарозной РНКазы.
— Степень амплификации ДНКазы I, концентрация 1 Ед / мкл. Thermo Fisher Scientific.
— Система синтеза первой цепи SuperScript TM IV.Thermo Fisher Scientific.
— Универсальный набор зондовых библиотек, человек с зондами №1 — №90. Рош (Велвин-Гарден-Сити, Великобритания).
— Праймеры для qRT-PCR, разработанные для конкретных целей. Размеры праймеров составляли от 18 до 20 п.н., продукты амплификации короче 110 п.н., конечная использованная концентрация составляла 250 нМ для каждого праймера (таблица 1).
— Мастер микс датчика LightCycler ® 480. Рош.
— Системный прибор LightCycler ® 480. Рош.
Таблица 1. Последовательности праймеров и зонды, использованные в этом исследовании.
Протеомика
— Центробежные фильтровальные установки. Amicon ® ultra-0,5 мл, 3К. Merck Millipore (Таллагрин, Ирландия).
— Vivaspin ® Мембрана для ультрафильтрации из полиэфирсульфона, 500 кДа, MWCO. GE Healthcare.
— Муравьиная кислота.
— UHQ (сверхвысокое качество)> 18 МОм воды.
— 50 мМ бикарбонат аммония (AMBIC).
— 200 мМ дитиотреитол (DTT) и 30 мг / мл AMBIC (раствор DTT).
— 200 мМ йодацетамида (IAA) плюс 36 мг / мл AMBIC (раствор IAA).
— Ацетонитрил (ACN).
— Раствор A: 0,1% трифторуксусная кислота (TFA) в воде UHQ.
— Раствор B: 0,1% TFA, 70% ACN в воде UHQ.
— Трипсин (Кат. № V511A). Промега.
— Q Exactive TM Гибридная четырехполюсная орбитальная ловушка. Thermo Fisher Scientific.
— UltiMate TM 3000 RSLnano система жидкостной хроматографии (ЖХ). Thermo Fisher Scientific.
— Ионный источник EASY-Spray TM . Thermo Fisher Scientific.
— Acclaim TM Pepmap TM 100 колонок C18 LC (каталожный номер 160454). Диаметр 300 мкм × длина 5 мм, размер частиц 5 мкм. Thermo Fisher Scientific.
— Наноколонка PepMap TM RSLC C18, внутренний диаметр 75 мкм × 25 см. Thermo Fisher Scientific.
— Наконечники Millipore ZipTips C18. Сигма-Олдрич.
— Proteome Discover TM Программное обеспечение версии 1.4. Thermo Fisher Scientific.
— программный пакет Mascot Server версии 2.5. Matrix Science Inc. (Лондон, Великобритания).
Пошаговые процедуры
Приготовление инокулята
T. rubrum— T. rubrum культивировали на КПК в течение 7–10 дней при 30 ° C.
— Конидии выделяли путем добавления 3-5 мл стерильной воды на поверхность грибкового роста и соскоба L-образным распределителем клеток.
— Жидкость была собрана и помещена в пробирку на 15 мл и оставлена на 2 мин в вертикальном положении.Супернатант собирали, и клетки трижды промывали PBS.
— Конечная плотность клеток была определена путем подсчета с помощью гемоцитометра и скорректирована для инфицирования эксплантата кожи 1 × 10 6 конидий в 10 мкл, что было подтверждено подсчетом жизнеспособных клеток (колониеобразующих единиц) на КПК.
Создание человеческой кожи
ex vivo Модель и грибковая инфекция— Эксплантаты кожи грудного и брюшного происхождения были получены из Tissue Solutions ® (рис. 1A).
— Человеческие ткани, предоставленные этой компанией, получены в соответствии с юридическими и этическими требованиями страны сбора, с этического одобрения и анонимного согласия донора или ближайшего родственника. Tissue Solutions ® также соответствует требованиям Управления по тканям Соединенного Королевства (HTA) в отношении импорта тканей.
— Эксплантаты транспортировали и отправляли в холодильных установках на 4 ° C и поддерживали при 4 ° C до обработки, которая произошла в течение 36 часов после операции.
— В DMEM добавляли 1% об. / Об. Антибиотиков (пенициллин и стрептомицин) и 10% HI-FBS.
— Кожу промывали DMEM и держали влажной в чашке Петри с той же средой (рис. 1B). Эксплант был разрезан на кусочки размером 1 см 2 ткани. Поверхность каждого куска размером 1 см 2 была осторожно ранена с помощью иглы, не пересекая всю толщину кожи. Поверхность кожи прокалывали иглой несколько раз (6–10).
— После ранения каждый кусочек кожи помещали в отдельные лунки 6-луночного планшета.Межфазную фазу воздух-жидкость поддерживали добавлением 1 мл дополненной среды DMEM. Добавленный объем среды покрыл только дерму, избегая контакта с эпидермисом (рис. 1C).
— Эксплант был инокулирован путем нанесения 10 мкл конидиальной суспензии T. rubrum непосредственно на эпидермис, избегая протекания и контакта с дермой и окружающей средой. Всегда включались контроли только с кожей (неинфицированные).
— Эксплантаты инкубировали при 37 ° C в 5% CO 2 в инкубаторе CO 2 .Среду меняли каждые 24 ч, израсходованную среду сохраняли в пробирках на 2 мл для последующего анализа. Восстановленные среды хранили при -80 ° C. Когда культуральную среду меняли, следили за тем, чтобы свежая среда не касалась поверхности эксплантата кожи, обеспечивая контакт кожи с воздухом только во время эксперимента.
— Образцы кожи также были извлечены в чашку Петри в разные моменты времени. Перед обработкой образцов кожи их макроскопический вид оценивали на глаз, а изображения получали с помощью стереомикроскопа Stemi 2000-c.
— Восстановленные образцы были затем обработаны в зависимости от дальнейших запланированных анализов и испытаний, которые описаны в следующих шагах.
— Ткань для гистологических тестов помещали в формы, а затем заливали в состав ОКТ и быстро замораживали с помощью сухого льда и изопентана. Эти образцы можно хранить при -20 ° C для немедленного анализа или при -80 ° C для длительного хранения.
— Для SEM ткань фиксировали в буфере глутаральдегида (2,5% глутарового альдегида в 0,1 М какодилате) в течение ночи при 4 ° C.Эти образцы необходимо обработать сразу после их извлечения.
— Ткань для экстракции РНК была разрезана на более мелкие части и помещена в РНК , позже ® для последующей экстракции РНК. Эти образцы могут храниться при разных температурах в зависимости от потребностей исследователя. Восстановленные ткани хранили при -80 ° C для более длительного хранения или -20 ° C для более быстрого использования.
— Эксперименты для каждого состояния повторяли не менее трех раз с использованием кожи разных людей-доноров.
Рисунок 1. Кожа человека ex vivo модель . (A) Эксплантаты кожи были получены от третьей стороны. Перед применением жировой слой отбрасывали. (B) Кожные эксплантаты разрезали на кусочки размером 1 см 2 . Осколки были ранены иглой. (C) Кусочки кожи поддерживали в культуральной среде, поддерживая межфазную фазу воздух-жидкость. Среда покрывала только дерму, избегая контакта с эпидермисом.
Гистологический анализ
— Для гистологического подтверждения грибковой инфекции срезы ткани толщиной 6 мкм, вырезанные из замороженных блоков OCT с использованием криостата, помещали на предметные стекла, покрытые поли- L -лизином, и фиксировали 4% параформальдегидом без метанола в PBS в течение 20 мин. перед дальнейшим окрашиванием.
Модифицированный метенамин серебра (GMS) с окрашиванием начальным раствором периодической кислоты (PAS)
— Срезы окрашивали модифицированным красителем GMS (HT-100) в соответствии с инструкциями производителя.
— Предметные стекла контрастировали либо раствором гематоксилина, либо 0,2% (мас. / Об.) Светло-зеленым желтоватым раствором SF в течение 1-2 мин.
— Предметные стекла были обезвожены в градиенте этанола с 30-секундной инкубацией при каждой концентрации (70%, 80%, 90%, 100%, 100%).
— Очистку проводили дважды с использованием гистологического очищающего соединения в течение 30 с.
— Слайды закрепляли с помощью состава HistoMount TM , накладывали покровное стекло и оставляли сушиться на ночь.
— Изображения были получены с помощью цифрового сканера Axio scan.Z1 с использованием окуляра 10 ×.
Калькофлуор Белый (CFW)
— Срезы также окрашивали CFW.
— Предметные стекла инкубировали в 10% (мас. / Об.) КОН в течение 15 минут, трижды промывали PBS и сушили на воздухе перед окрашиванием 1% (мас. / Об.) CFW в течение 15 минут, затем снова трижды промывали PBS. .
— Предметные стекла окрашивали раствором иодида пропидия (PI) (1 мкг / мл в PBS) в течение 15 минут, трижды промывали PBS и позволяли высохнуть в темноте при комнатной температуре (RT) в течение 30 минут или до высыхания.
— Одна капля монтажной среды Vectashield ® была помещена на окрашенный участок и покровное стекло использовалось для равномерного распределения монтажной среды.
— Изображения были получены с помощью конфокального микроскопа DeltaVision с увеличением 10 ×.
Жизнеспособность тканей
— Жизнеспособность клеток кожной ткани оценивалась в незараженных тканях с использованием системы TUNEL.При обнаружении фрагментированной ДНК, то есть апоптозных или умирающих клеток, следовали инструкциям производителя.
— Предметные стекла окрашивали раствором PI (1 мкг / мл в PBS в течение 15 минут), трижды промывали PBS и позволяли высохнуть в темноте при комнатной температуре в течение 30 минут или до высыхания.
— После сушки слайды помещали в Vectashield ® плюс DAPI для окрашивания ядер.
— Изображения были получены с помощью конфокального микроскопа DeltaVision с увеличением 10 ×.
— Уровень апоптоза оценивали путем деления количества клеток, окрашенных системой TUNEL, на общее количество клеток, окрашенных с помощью трех методов окрашивания (DAPI, йодид пропидия и TUNEL).Затем уровни сравнивали с незараженной кожей через 24 часа с использованием программного обеспечения Prism 7 v.7.1 с использованием однофакторного дисперсионного анализа и апостериорного теста Даннета. Статистически значимым считалось значение p <0,05.
Сканирующая электронная микроскопия (СЭМ)
— Ткани немедленно фиксировали в 2,5% глутаровом альдегиде в 0,1 М буфере какодилата натрия, pH 7,2 и инкубировали в течение ночи при 4 ° C.
— Образцы промывали 0,1 М натрий-какодилатным буфером (pH 7.2–7.4) дважды по 5 мин.
— Ткани обрабатывали 1% тетроксидом осмия в дистиллированной воде в течение 1 часа, затем трижды промывали натриевым какодилатным буфером в течение 5 минут.
— Образцы дегидратировали в градиенте этанола (70%, 80%, 90%), при этом образцы инкубировали в течение 10 минут при каждой концентрации.
— Образцы трижды инкубировали в 100% этаноле в течение 10 мин.
— Образцы инкубировали в течение 3 минут в HMDS и помещали в эксикатор на 25 минут для удаления любых водных загрязнений.
— Образцы наблюдались с помощью сканирующего электронного микроскопа Zeiss TM EVO MA10 (несколько различных увеличений).
Количественная ОТ-ПЦР для анализа экспрессии генов человека
Экстракция РНК и производство кДНК
— РНК позже раствор ® был отброшен, а ткань промыта стерильной водой.
— Ткань разрезали на мельчайшие кусочки, помещали в ступку и измельчали. Жидкий азот использовали для быстрой заморозки ткани и снова измельчали до тех пор, пока маленькие кусочки не превратились в порошок.
— Порошок ткани помещали в пробирки на 2 мл, содержащие шарики матрицы D, и добавляли 1 мл TRIzol TM . Пробирки помещали в прибор FastPrep TM и обрабатывали со скоростью 6 м / с в течение трех-пяти циклов по 40 с с интервалами 5 мин. Пробирки инкубировали на льду в течение интервалов.
— Пробирки центрифугировали при 14000 об / мин в течение 5 минут при 4 ° C и супернатант переносили в свежие 2 мл микроцентрифужные пробирки.
— в каждую пробирку добавляли 0,2 мл хлороформа / изоамилового спирта (49: 1).Пробирки встряхивали в течение 10 с, затем охлаждали на льду в течение 15 мин.
— Пробирки центрифугировали 20 мин при 10000 об / мин при 4 ° C.
— Водный слой переносили в новую пробирку на 2 мл. Органический (розовый) слой также сохраняли для последующей экстракции белка (см. Раздел «Протеомика и анализ дробовика»).
— 1 мл изопропанола добавляли к выделенному водному слою и оставляли на ночь при -20 ° C для осаждения РНК.
— Образцы центрифугировали в течение 20 мин при 10 000 об / мин при 4 ° C и супернатант отбрасывали.Осажденная РНК представляет собой гелеобразный осадок.
— Произведено второе осаждение. Осадок РНК ресуспендировали в 0,3 мл тризола, затем переносили в пробирку на 1,5 мл. Добавляли изопропанол (0,3 мл) и пробирки инкубировали не менее 30 мин при -20 ° C.
— Образец центрифугировали в течение 10 мин при 10 000 об / мин при 4 ° C и супернатант отбрасывали.
— Осадок растворяли в 1 мл 75% этанола и встряхивали в течение нескольких секунд, затем инкубировали в течение 10-15 минут при комнатной температуре.
— Образец центрифугировали в течение 5 минут при 10 000 об / мин при 4 ° C, супернатант удаляли и осадок сушили на воздухе при комнатной температуре в течение 5–10 минут. Этап очистки с использованием 1 мл 75% этанола повторяли два раза перед тем, как осадок РНК ресуспендировали в 50–100 мкл стерильной воды, обработанной DEPC, а затем инкубировали при 60 ° C в течение 15 мин.
— Выход и чистота РНК оценивали с помощью NanoDrop TM , и образцы хранили при -80 ° C.
— Целостность / качество РНК оценивали путем обработки 5 мкл общей РНК на 1% неденатурирующем агарозном геле TBE в 1X буфере TBE.Перед использованием резервуары очищали с помощью RNaseZap TM .
— Для получения кДНК образцы РНК (1 мкг) обрабатывали ДНКазой I, затем проводили обратную транскрипцию с использованием системы синтеза первой цепи SuperScript TM IV, следуя инструкциям производителя.
Дизайн, реакции и условия анализа количественной ПЦР с двумя датчиками гидролиза
— Праймеры и анализы, охватывающие интроны, были разработаны с использованием Roche’s Universal Probe Library Assay Design Center для пяти различных генов-мишеней, которые, как известно, экспрессируются в коже и во время T.rubrum (таблица 1).
— Один референсный ген ( B2M для β2-микроглобулина) был также разработан (Lossos et al., 2003) с использованием веб-инструмента Eurogentec для модификаций для добавления необходимого зонда (Таблица 1).
— Реакции qRT-PCR объемом 10 мкл проводили в планшетах LightCycler 480 с использованием мастер-микса зонда LightCycler 480 в соответствии с инструкциями производителя (таблица 2).
— Реакции проводили в LightCycler 480. Установки были следующими: предварительная инкубация при 95 ° C в течение 10 мин (скорость изменения 4.8 ° C / с), один цикл, 55 циклов фазы амплификации с денатурацией при 95 ° C в течение 10 с (скорость изменения 4,8 ° C / с), отжиг при 60 ° C в течение 30 с (скорость изменения 2,5 ° C / с ), удлинение 72 ° C в течение 1 с (скорость изменения 4,8 ° C / с) и, наконец, один цикл фазы охлаждения при 40 ° C в течение 30 секунд (скорость изменения 2,5 ° C / с).
— Анализы на каждом образце кДНК проводили в трех экземплярах. Была проведена относительная количественная оценка. Результаты, полученные для каждого целевого гена, нормализовали по уровням экспрессии гена β2-микроглобулина. Результаты неинфицированной кожи использовали в качестве отрицательного контроля для инфекции.Результаты были проанализированы с использованием метода 2 -ΔΔC T (Livak and Schmittgen, 2001).
— Статистический анализ проводился с использованием критерия Стьюдента t или критерия Манна-Уитни в зависимости от распределения данных. Статистически значимым считалось значение p <0,05. Графики построены с использованием программного обеспечения Prism 7 (GraphPad, La Jolla, CA, США).
Таблица 2. реакций qRT-PCR.
Протеомика и анализ дробовика
Извлечение белков человека из кожи
— 300 мкл 100% этанола добавляли к выделенной органической фазе (розовый слой) после экстракции тканевой РНК, упомянутой в разделе «Количественная ОТ-ПЦР для анализа экспрессии генов человека». Пробирку перемешивали переворачиванием и оставляли на RT в течение 3 мин.
— Образцы центрифугировали при 3000 об / мин в течение 5 минут при 4 ° C.
— Супернатанты разделяли на две части и переносили в новые пробирки на 1,5 мл с добавлением 800 мкл изопропанола, а затем инкубировали в течение ночи при -20 ° C для осаждения белков.
— Образцы центрифугировали при 12000 об / мин в течение 10 мин при 4 ° C.
— Осадок промывали холодным 0,3 М гидрохлоридом гуанидина в 95% этаноле и оставляли на 20 мин при комнатной температуре.
— Образцы центрифугировали при 7500 об / мин в течение 5 минут при 4 ° C, а затем супернатант удаляли.Промывку повторили.
— Половина образца белка хранилась при -20 ° C (в 0,3 М гидрохлориде гуанидина в 95% этаноле) для расщепления трипсином и дальнейшего анализа с помощью ЖХ-МС / МС в Aberdeen Proteomics Facility.
— Другую половину образца белка промывали 95% этанолом и центрифугировали при 7500 об / мин в течение 5 минут при 4 ° C, супернатант удаляли и осадок сушили на воздухе.
— Осадок ресуспендировали в 100 мкл буфера для ресолюбилизации (1% мас. / Об. DTT, 2 M тиомочевина, 7 M мочевина и 4% мас. / Об. CHAPS, 2% об. Амфолитов-носителей, 10 мМ Pefabloc ® SC ингибитор протеиназы) и концентрацию белка, определяемую анализом Кумасси.
Извлечение белков из среды тканевой культуры
— Восстановленная / отработанная среда фильтровалась с использованием центробежных фильтров 50K Amicon ® Ultra-15 (Merck) и концентрировалась с помощью центробежных фильтров 3K Amicon ® Ultra-15 до тех пор, пока не оставалось 200 мкл. Этот шаг был необходим для удаления большинства белков сыворотки (в основном альбумина из HI-FBS, используемого во время культивирования (белки менее 50 кДа).
— Четыре временных точки (24 часа, 2, 6 и 8 дней) в течение T.rubrum , плюс отрицательный контроль, соответствующий используемой модифицированной среде, и среда из незараженной кожи.
Переваривание трипсина
— Концентрированные супернатанты и экстрагированные белки из ткани были расщеплены трипсином. Белки, извлеченные из ткани и хранящиеся при -20 ° C в гуанидин гидрохлориде (процедура, описанная в разделе «Извлечение белков человека из кожи»), центрифугировали при 7500 об / мин в течение 5 минут при 4 ° C и супернатант отбрасывали. Оставшийся осадок разводили в 100 мкл AMBIC.
— Количество используемых тканевых белков составляло 10 мкг на образец, разведенный в 100 мкл AMBIC. Для концентрированных супернатантов использовали 100 мкл на образец.
— раствор DTT (2 мкл) добавляли к каждому образцу белка (конечная концентрация 2 мМ) и образцы инкубировали в течение 25 мин при 60 ° C.
— Раствор ИУК (4 мкл) добавляли к каждому образцу (конечная концентрация 4 мМ), затем пробирку инкубировали в течение 30 мин при комнатной температуре в темноте.
— Разбавьте образцы в четыре раза AMBIC (конечная концентрация мочевины 2 M).
— Трипсин добавляли в соотношении 1: 100, и пробирки инкубировали в течение 16 часов при 37 ° C.
— Реакции замораживали при -70 ° C, сушили центрифугированием в вакууме, затем ресуспендировали в 40 мкл раствора A, перемешивали на вортексе в течение 10 минут, центрифугировали при 14000 об / мин.
— Пептиды были обессолены перед ЖХ-МС / МС с использованием C18 Ziptips. Сначала материал C18 кондиционировали 20 мкл ACN, затем промывали C18 20 мкл раствора B. Затем C18 уравновешивали 20 мкл раствора A. Каждый образец загружали в C18 и медленно отсасывали 20 мкл образца через наконечник 10 раз. .Образец дважды промывали 20 мкл раствора А. Наконец, образец элюировали 5 мкл раствора В. Образцы сушили центрифугированием в вакууме.
Жидкостная хроматография-масс-спектрометрия (ЖХ-МС / МС)
— Идентификация белка была проведена в лаборатории Aberdeen Proteomics Facility.
— Образцы повторно растворяли в 10 мкл 0,1% муравьиной кислоты в воде UHQ для получения концентраций пептида от 50 до 1000 фмоль / мкл.
— Жидкостная хроматография была выполнена с использованием системы нано-жидкостной хроматографии UltiMate TM 3000 RSL, сконфигурированной для предварительного концентрирования на наноколонке PepMap RSLC C18, установленной на ионном источнике EASY-Spray TM .
— Растворителем загрузочного насоса был UHQ вода: ацетонитрил: муравьиная кислота (98: 2: 0,1), и колонка работала со скоростью потока 10 мкл / мин.
— Растворителем А нанонасоса был UHQ вода: муравьиная кислота (100: 0,1).
— Растворителем В нанонасоса был ацетонитрил: вода UHQ: муравьиная кислота (80: 20: 0,1).
— Градиент ЖХ для программы «QC hemel v2.»
— Образцы вводили и переносили в предварительную колонку Pepmap TM 100 C18 300 мкм × 5 мм. Поток через предколонку переключили на нанонасос, и образец промыли обратно в аналитическую колонку и систему масс-спектрометрии (МС).
— Образцы были проанализированы методом восходящей протеомики с использованием полного сканирования MS / ddMS 2 (TopN) на платформе Q Exactive TM Hybrid Quadrupole Orbitrap с источником наноэлектрораспыления EASY-Spray TM .
— Настройки МС полного сканирования: разрешение 70 000, диапазон сканирования 375–1 750 м / з, максимальное IT 50 мс, цель АРУ 3e6. Параметры ddMS 2 : разрешение 17 500, изоляция 1,6 м / z, максимальное IT 100 мс и цель AGC 5e4. TopN было 10, что относится к количеству сканирований MS2.
— Сырые файлы были обработаны с помощью Proteome Discoverer v.14 для идентификации белков. Поиск в базе данных проводился с помощью Mascot server v 2.5 с использованием последовательностей белков Homo sapiens (база данных Swiss-Prot, версия 2017_01; 553474 последовательности, загружены 26.01.17, отфильтрованы по таксономии = Homo sapiens ).
— Параметры узла талисмана учитывают следующие динамические модификации во время переваривания трипсина: окисление (M) и карбамидометил (C). Уровень ложного обнаружения (FDR) был установлен на 0.01. Количественное определение белка выражалось как площадь под кривой.
— Четыре биологических повтора для каждого состояния ( T. rubrum, инфицированная и незараженная кожа) были проанализированы с помощью LC-MS / MS. Был включен пустой образец (только средний — без кожи человека). Белки, идентифицированные в холостом ходе, были исключены из анализа. Затем отбирали только белки, имеющие два или более идентифицированных пептида и два или более совпадения пептидного спектра (PSM). Наконец, белки, обнаруженные как минимум в двух из четырех проанализированных образцов для каждого условия, были включены для дальнейшего анализа онтологии генов (GO) с использованием онлайн-инструмента консорциума GO.
— Значения площади каждого белка усреднялись и сравнивались между условиями, инфицированной и незараженной кожей T. rubrum и, наконец, анализировались с использованием теста Стьюдента t или теста Манна-Уитни в зависимости от распределения данных a значение p <0,05 считалось статистически значимым.
Результаты
Подтверждение инфекции макроскопическим исследованием и микроскопическими доказательствами
После инокуляции кожи среду собирали и меняли каждые 24 часа.Макроскопический вид кожи тщательно наблюдали каждые 24 часа. Повреждение ткани инфицированной кожи было макроскопически видимым через 3 дня, при этом поражение увеличивалось с каждым днем, и через 10 дней оно было почти таким же большим, как кусок кожи размером 1 см 2 (рис. 2).
Рисунок 2. Макроскопическое подтверждение инфекции. (A) Кожа без инфекции. (B) T. rubrum инфицировал ex vivo кожи человека. Масштабная линейка на каждом рисунке соответствует 1 мм.
Наблюдаемое поражение кожи имело ожидаемый макроскопический вид, наблюдаемый в клинике, где поражения стригущего лишая или опоясывающего лишая описываются как плоская чешуйчатая область на коже, красная, с расширяющимися границами, образующими кольцо с прозрачной, чешуйчатой или внутренней частью. красные шишки. Такой же макроскопический вид наблюдали во всех биологических повторностях (кожа от разных доноров). Это подтверждает воспроизводимость модели.
Устранение неполадок 1 : Сообщалось, что конидии можно использовать в течение месяца после приготовления посевного материала при хранении при 4 ° C (Faway et al., 2017). Однако при использовании более старых конидий требуется больше времени, чтобы увидеть на глаз макроскопические признаки инфекции.
Устранение неисправностей 2 : Манипуляции с кожей могут привести к ее загрязнению. Использование раствора Chemgene (5%) и этанола (70%) для очистки рабочих поверхностей снижает вероятность загрязнения. Все хирургические инструменты необходимо стерилизовать перед использованием и очищать раствором Chemgene после использования.
Инфекции, вызываемые T. rubrum , будут поверхностными, то есть ограниченными эпидермисом.Модифицированное окрашивание серебром и CFW подтвердили присутствие гиф на поверхности кожи (рис. 3). Кроме того, было замечено, что эпидермис отделился от дермы. Этот вывод был подтвержден на нескольких слайдах и в повторах от разных доноров.
Рисунок 3. Гистологическое подтверждение и сканирующая электронная микроскопия через 10 дней. Неинфицированная кожа (A, C, E) сравнивалась с T. rubrum -инфицированной кожей (B, D, F) . Панели (A, B) показывают окрашивание серебром и контрастное окрашивание гематоксилином.Пунктирная стрелка указывает гиф T. rubrum . Сплошные стрелки обозначают эпидермис. Панели (C, D) показывают окрашивание CFW (синий), идентифицирующее хитин, следовательно, клеток T. rubrum . Окрашивание ядер эпидермиса пропидием йодидом (красный). Панели (E, F) показывают изображения SEM, где масштабная линейка составляет 10 мкм.
Поиск и устранение неисправностей 3 : При покрытии образцов компаундом для ОКТ необходимо избегать образования пузырьков. Состав для ОКТ должен покрывать ткань, чтобы уменьшить повреждение из-за мгновенного замораживания.
Устранение неполадок 4 : Коричневый цвет гиф после окрашивания серебром может быть светлее, чем ожидалось при окрашивании гематоксилином. Этого можно избежать, если в качестве контрастного красителя использовали светло-зеленый SF, или когда окрашивание проводилось классическим окрашиванием серебром с использованием 10% хромовой кислоты вместо PAS, или если образцы предварительно нагревали перед добавлением PAS.
The DeadEnd TM флуорометрический анализ системы TUNEL измеряет фрагментацию ядерной ДНК как маркер апоптоза и жизнеспособности клеток.Долю апоптоза оценивали путем деления количества клеток, окрашенных системой TUNEL, на общее количество окрашенных клеток. Затем сравнивали пропорции между тремя проанализированными состояниями: незараженная кожа через 24 часа, незараженная кожа через 14 часов и кожа, инфицированная T. rubrum. Макроскопический вид незараженной кожи после инкубации не отличался. Чтобы подтвердить присутствие грибков на поверхности инфицированной кожи, было проведено прямое наблюдение с помощью SEM.Кроме того, по данным SEM, эпидермальные клетки выглядели приподнятыми из-за присутствия грибов на поверхности кожи (рисунки 3E, F).
На 10-й день поверхность незараженной кожи была очень похожа на внешний вид кожи на 1-й день. Жизнеспособность кожи была подтверждена с помощью системы TUNEL. Через 14 дней не наблюдалось увеличения количества апоптотических клеток по сравнению с кожей, инкубированной в течение 24 часов (рис. 4). Доля апоптоза в инфицированной коже через 10 дней значительно отличалась от неинфицированной кожи через 24 часа (0.6 ± 0,07 против 0,002 ± 0,004, p <0,0001) и 14 дней (0,6 ± 0,07 против 0,04 ± 0,009, p <0,0001) (Рисунок 4).
Рис. 4. Жизнеспособность клеток , измеренная системой TUNEL. (A) Неинфицированная кожа через 24 часа. (B) Неинфицированная кожа через 14 дней. (C) T. rubrum -инфицированная кожа через 10 дней. На всех трех изображениях окрашивание йодидом пропидия (красно-розовый), окрашивание DAPI (синий), чтобы показать неапоптотические клетки, оба красителя связывают ДНК.Апоптотические клетки окрашены в зеленый цвет с помощью системы TUNEL для идентификации фрагментированной ДНК. Долю апоптоза оценивали путем деления количества апоптотических клеток (зеленых клеток) на общее количество клеток (зеленые и красно-розовые клетки). Результаты на графике представляют четыре биологических повтора ( n = 4), с тремя секциями, анализируемыми на секцию. Данные сравнивали с помощью однофакторного дисперсионного анализа. p = 0,0002, затем апостериорного теста Даннета , где ∗∗∗ представляет p <0.0001.
Экспрессия гена
Кератиноциты продуцируют несколько цитокинов, включая IL8 (CXCL8) и IL18 (член семейства IL-1β) (Pasparakis et al., 2014). Рецептор IL6 также экспрессируется в пролиферативных кератиноцитах (Hänel et al., 2013). Трансформирующий фактор роста бета 1 (TGFB1) является основным регулятором биологии дендритных клеток в коже, а CCL20 является мощным хемокином для лимфоцитов и дендритных клеток (Nedoszytko et al., 2014). Экспрессия всех пяти генов, кодирующих цитокины, была подтверждена в неинфицированной и инфицированной коже через 14 и 10 дней инкубации, соответственно (четыре экспериментальных образца для каждого состояния, каждый измерен в трех экземплярах).Кратность изменения в инфицированных образцах оценивали с использованием неинфицированной кожи того же донора, что и контрольный образец, после нормализации с использованием гена β2-микроглобулина в качестве контрольного гена.
Экспрессия IL8 (двукратная, p = 0,05) и CCL20 (двукратная, p = 0,05) была увеличена во время инфицирования T. rubrum , но не было различий в экспрессии IL6R и TGFB1 между инфицированной и незараженной кожей (рис. 5).Наконец, экспрессия IL18 была снижена (2E5 раз, p = 0,03) в инфицированной коже по сравнению с неинфицированной кожей (фиг. 5).
Рисунок 5. Экспрессия гена эксплантата кожи, оцененная через 10 дней после инфицирования. Относительную экспрессию оценивали с использованием неинфицированной кожи того же донора в качестве эталонного образца для нормализации с геном B2M. N = 4 биологических повтора, проанализированных в трех повторах. Данные представлены в виде медианы и IQR, проанализированных с помощью теста Манна – Уитни U .
Устранение неисправностей 5 : Во время экстракции РНК кожи необходимо разрезать, раздробить или измельчить кожу на мелкие кусочки, поскольку это увеличивает количество РНК, экстрагированной из грибковых и человеческих клеток в образце.
Устранение неисправностей 6 : При использовании TRIzol для экстракции РНК необходимо дважды осаждать РНК для повышения чистоты РНК. Промывку 75% этанолом следует проводить дважды или трижды, чтобы избежать загрязнения белками и фенолом.
Протеомика
Четыре биологические повторы для каждого условия ( T.rubrum инфицированную и незараженную кожу через 10 дней инкубации для образцов ткани и через 1, 2, 4 и 8 дней инкубации для супернатантов) анализировали с помощью LC-MS / MS. Идентифицированные белки были проверены и отобраны для дальнейшего анализа онтологии генов (GO) с использованием онлайн-инструмента консорциума GO (см. Текстовую сноску 3).
Количество белков, идентифицированных в ткани с помощью дробового анализа, было выше, чем соответствующее количество, идентифицированное в восстановленной среде / супернатанте. В ткани был обнаружен в общей сложности 2451 белок, но только 1229 белков удовлетворяли критериям включения для дальнейшего анализа.Между тем, в отфильтрованных и извлеченных супернатантах было обнаружено в общей сложности 523 белка, но только 201 соответствовал критериям включения для дальнейшего анализа. После анализа генной онтологии белков, обнаруженных в тканях и супернатантах, наиболее важными биологическими процессами в обоих случаях были идентифицированы клеточные процессы и метаболические процессы. Ответ на стимул был третьим по важности процессом в супернатантах (рис. 6).
Рисунок 6. Процент биологических процессов ГО, происходящих в течение Т.rubrum инфекция. (A) Биологический процесс на основе белков, идентифицированных в ткани (процент совпадений гена по сравнению с общим числом совпадений процесса, совпадения = 1553). (B) Биологический процесс из белков, идентифицированных в супернатантах (процент совпадений гена по сравнению с общим числом совпадений процесса, совпадения = 302).
Специфические белки, участвующие в пролиферации и дифференцировке кератиноцитов, были идентифицированы в неинфицированной коже. Белки, участвующие в усилении дифференцировки (Кератин 9, пятикратный, p = 0.017) и поздней стадии дифференцировки (каспаза 14, -трёхкратная, p = 0,015) значительно различались в инфицированной коже (фигура 7). Уровни отдельных белков, обнаруженных в инфицированной коже, были нормализованы по отношению к уровням, обнаруженным в незараженной коже, и сравнивались с помощью теста Student t .
Рис. 7. Доказательства продолжающегося размножения в коже во время инфекции T. rubrum . В таблице показано кратное изменение уровней белка в коже, инфицированной T.rubrum относительно уровней, обнаруженных в незараженной коже. Кератины, выделенные курсивом, обнаруживаются в эпителиальных клетках. ∗ Студент t — результаты теста, где p <0,05 является статистически значимым. Кератины, участвующие в пролиферации и дифференцировке, показаны цветным текстом. На рисунке показаны функции и локализация кератинов в эпидермисе.
Устранение неполадок 7 : Извлечение РНК и белков из одного и того же образца кожи увеличивает время, необходимое для его обработки.Оба процесса экстракции можно остановить на стадии осаждения и хранить при -20 ° C не менее года, что позволяет завершить процесс позже.
Устранение неполадок 8 : На чувствительность Q-Exactive могут влиять белки в буфере для ресолюбилизации. Поэтому мы разделили осадок белка пополам, что позволило нам количественно определить количество белков в буфере для ресолюбилизации и оценить количество белков в другой половине, которые должны быть переварены трипсином.
Устранение неполадок 9: Несмотря на то, что фильтрация проводилась перед ЖХ-МС / МС, чтобы избежать большей части сывороточного альбумина из FCS, альбумин все же был идентифицирован с помощью Q-Exactive.Во время биоинформатического анализа из него исключили альбумин, предположив, что он является загрязняющим веществом. Важно иметь образец среды с добавками для использования в качестве холостого контроля и анализировать его с помощью ЖХ-МС / МС перед анализом супернатантов, полученных в экспериментах с кожей, во избежание смещения из-за белков из FCS.
Обсуждение
В этом исследовании мы описываем методологию изучения SFI с использованием эксплантатов кожи человека. Этот метод позволяет воспроизводить результаты, несмотря на то, что кожа получена от разных доноров.Использование этой модели ex vivo дает более релевантные результаты, отражающие то, что происходит in vivo по сравнению с другими моделями. Было показано, что на моделях эпидермиса животных наблюдаются другие воспалительные процессы, чем у людей. Эти различия зависят, во-первых, от различий в метаболизме и анатомии кожи человека и животных. Эпидермис и дерма мышей не такие многослойные и толстые, как кожа человека (Groeber et al., 2011). Кроме того, процесс заживления ран у мышей происходит быстрее и не оставляет рубцов.Кроме того, некоторые подтипы дендритных клеток мышей не обнаруживаются в коже человека (Gudjonsson et al., 2007). Наконец, реконструированные модели эпидермиса мышей показали, что экспрессия генов цитокинов, в частности IL22 и IL17R , в эпидермисе человека выше, чем в реконструированном эпидермисе мышей (Pohin et al., 2017).
Суррогаты для кожи человека, такие как однослойная культура кератиноцитов человека, слой рогового слоя и реконструированный эпидермис, также имеют некоторые ограничения, которых можно избежать с помощью эксплантатов кожи.Хотя реконструированный эпидермис обладает почти всеми преимуществами эксплантатов кожи полной толщины, в большинстве этих моделей отсутствуют дендритные клетки и меланоциты (Groeber et al., 2011). Культуры монослоя кератиноцитов и слой роговицы плохо дифференцируются или не дифференцируются, соответственно, что приводит к более низкому количеству экспрессируемых генов или различным паттернам экспрессии генов по сравнению с реконструированным эпидермисом и / или нормальным эпидермисом. Некоторые из генов, которые не экспрессируются или имеют разные паттерны экспрессии, включают ICAM1, , цитокератины, десмоколлин3 и лорикрин (Bernard et al., 2002; Мехул и др., 2004; Rechavi et al., 2006). Кроме того, использование полной толщины ex vivo кожи человека и реконструированного эпидермиса позволяет изучать местные лекарственные средства, местные эффекты антисептиков и биопленки.
Национальный центр по замене, усовершенствованию и сокращению количества животных в исследованиях (NC3Rs) по социальным и этическим причинам выступает за сокращение и адаптацию использования животных в фармацевтических, химических, пестицидных и косметических исследованиях (Burden et al., 2015; Прескотт и Лидстер, 2017). Использование эксплантатов кожи человека в исследовании SFI представляет собой альтернативу использованию на животных.
Другие потенциальные преимущества модели кожи человека ex vivo для исследования SFI были показаны Poyntner et al. (2016). Эта группа описала и улучшила количество аннотированных генов, кодирующих белок в транскриптоме Exophiala dermatitidis (Poyntner et al., 2016), и протестировала мутант поликетидсинтазы ( PKS1 ) E.dermatitidis , чтобы оценить роль меланина во время кожной инфекции (Poyntner et al., 2018).
Проблема использования кожных эксплантатов в исследованиях связана с продолжительностью жизни кожной ткани. В некоторых исследованиях сообщается о более низкой продолжительности жизни и высокой скорости апоптоза кожных эксплантатов, например, инкубация в течение 5–6 дней при 37 ° C (Kleszczyński and Fischer, 2012). Однако в этих исследованиях сыворотка не добавлялась в среду для роста кожи. Кроме того, инкубация полной толщины кожи (эпидермиса и дермы) улучшает пролиферативную способность кератиноцитов (Lee and Cho, 2005).Известно, что фибробласты в дерме играют роль в поддержании пролиферации эпидермиса дольше, чем модели, которые используют только эпидермис или монослои кератиноцитов (Groeber et al., 2011). Мы не наблюдали различий в уровнях апоптоза в неинфицированной кожной ткани через 24 часа, 10 и 14 дней инкубации, подтверждая, что кожа остается жизнеспособной в течение 10 и 14 дней при 37 ° C.
В этом исследовании было обнаружено больше доказательств жизнеспособности и пролиферативной способности ткани после 10 дней инкубации.Доказательства экспрессии IL6R были обнаружены в неинфицированной коже, которая не отличалась от инфицированной кожи, наряду с присутствием филаггрина и цитокератинов K1, K10, K5, K14, K15 в протеомном анализе. Кератины — это филаменты цитоскелета, фундаментальные для структурной стабильности и механики многослойного эпидермиса. Экспрессия кератина различна во время клеточной дифференцировки. Кератины K5 / K14 / K15 являются основными кератинами, экспрессируемыми в недифференцированном базальном слое кератиноцитов (содержащем стволовые клетки), между тем, K1 / K10 являются основными кератинами в процессе дифференцировки кератиноцитов, а K2, филаггрин и каспаза 14 являются маркерами для терминальный процесс дифференцировки, ороговение и самые внешние слои эпидермиса (Denecker et al., 2008; Молл и др., 2008). В этом исследовании были обнаружены все эти кератины, что свидетельствует о всех стадиях пролиферации и дифференцировки в неинфицированной коже.
Во время инфекции, вызванной T. rubrum , были значительно затронуты три белка, участвующих в пролиферации и дифференцировке кератиноцитов (K9, K18 имели более высокие уровни, а уровни каспазы 14 были снижены). Характер этих белков во время кожной инфекции T. rubrum добавил доказательства индукции апоптоза, который наблюдали с использованием системы TUNEL в гистологических срезах.Каспаза 14, в отличие от других каспаз, не активируется апоптозом. Более низкие уровни этого белка во время грибковой инфекции отражают более низкую дифференцировку кератиноцитов (Rendl et al., 2002). Кроме того, в инфицированной коже были обнаружены более высокие уровни K18, которые не были обнаружены в незараженной коже. K18 обычно не обнаруживается в нормальной здоровой коже, однако его идентификация при воспалительных состояниях была описана как хороший маркер раннего апоптоза (Caulín et al., 1997).
В нашей модели грибковая инфекция была инициирована после повреждения эпидермиса иглой.Однако в будущем также можно будет исследовать, могут ли грибковые инфекции инициировать успешные инфекции, не повреждая поверхность кожи, а также влияют ли различные методы повреждения на взаимодействия хозяин-патоген и, в конечном итоге, на развитие успешной инфекции. Другие методы повреждения, которые можно исследовать на предмет их влияния на грибковую инфекцию, включают истирание (Gao et al., 2013), УФ-повреждение (Khalil, 2018), лазерное повреждение (Marquardt et al., 2015) или ожоги (Emanuelsson and Kratz, 1997).
Нейтрофилы, в отличие от Т-лимфоцитов и дендритных клеток, не являются нормальными резидентными клетками кожи, но рекрутируются и локально присутствуют в большом количестве через 24 часа после повреждения кожи (Gillitzer and Goebeler, 2001). Рекрутирование нейтрофилов не происходило или происходило не полностью для эксплантатов кожи, используемых в этих экспериментах, поскольку образцы кожи берутся у донора сразу после хирургического разреза кожи донора. Следовательно, модель, использованная в наших экспериментах, может рассматриваться как нейтропеническая кожа.Однако модель все еще может быть использована для оценки передачи сигналов рекрутирования нейтрофилов. Доказательства этого процесса наблюдались во время инфицирования T. rubrum , поскольку экспрессия гена CXCL8 была увеличена в ткани, что свидетельствует о попытках привлечь иммунные клетки к поврежденной ткани. Это один из недостатков этой модели, поскольку она моделирует только прямые взаимодействия и ответы клеток кожи с грибами. Отсутствие вовлечения кровотока и циркулирующих иммунных клеток ограничивает возможности моделирования взаимодействий между хозяином и патогеном; однако в будущем можно будет расширить модель, дополнив модель иммунными клетками, добавленными в культуральную среду (Kühbacher et al., 2016).
Дополнительные доказательства иммунного ответа на T. rubrum могут быть получены из наших результатов. Более высокая экспрессия CCL20 во время инфекции T. rubrum может отражать попытки рекрутировать Treg и дендритные клетки (Homey et al., 2000). Во время инфекции T. rubrum мы обнаружили, что IL18 имеет сниженную экспрессию, что может отражать то, что клетки TH 2 не задействуются или не возникает IFN-зависимого хронического воспаления (Nakanishi et al., 2001). Кроме того, низкая экспрессия IL18 может быть связана с деструктивным воздействием на эпидермис, наблюдаемым во время инфицирования T. rubrum в этом исследовании.
Основные недостатки использования кожных эксплантатов при исследовании SFI связаны с доступом к ткани и межиндивидуальными вариабельными реакциями на инфекцию. Доступ к кожным эксплантатам, использованным в этом проекте, осуществлялся через третье лицо. Другие варианты доступа к тканям могут быть через местные больницы. В первом варианте третье лицо помогает с этическим одобрением; однако стоимость эксплантатов выше, чем при получении местных тканей.Недостатками при подаче этического одобрения на месте может быть время, затрачиваемое на получение одобрения, и эти задержки следует учитывать при планировании исследований с использованием кожных эксплантатов.
В предыдущих отчетах и обзорах по использованию кожных эксплантатов для изучения различных заболеваний сообщалось о межиндивидуальной вариабельности среди доноров кожи (Bernard et al., 2002; Groeber et al., 2011). В нашем исследовании эта вариабельность была обнаружена только во время анализа экспрессии генов, когда повышенная или пониженная экспрессия конкретного гена была более очевидной для одного или двух биологических повторов, но с аналогичной тенденцией, наблюдаемой для остальных образцов.
Заключение
Протокол, представленный здесь, описывает методологию успешного моделирования кожи человека T. rubrum инфекции. Это воспроизводимая модель, которая представляет собой вариант изучения не только этой инфекции, но и других SFI, с получением более близких результатов к тем, которые мы обычно наблюдаем клинически. Важно отметить, что это согласуется с целями NC3R, заменяя использование животных в исследованиях.
Авторские взносы
DC-L участвовал в разработке экспериментов и выполнил эксперименты, описанные в рукописи, проанализировал данные и написал рукопись.КМ участвовал в управлении проектом, наблюдении и редактировании рукописи. DM является автором-корреспондентом, главным научным руководителем и участвовал в написании и редактировании рукописи.
Финансирование
Этот проект финансировался стратегической премией Wellcome Trust в области медицинской микологии и грибковой иммунологии 097377. Мы хотели бы поблагодарить Центр медицинских исследований по медицинской микологии при Университете Абердина (MR / N006364 / 1) за поддержку.
Заявление о конфликте интересов
Авторы заявляют, что исследование проводилось при отсутствии каких-либо коммерческих или финансовых отношений, которые могут быть истолкованы как потенциальный конфликт интересов.
Благодарности
Спасибо г-же Люсинде Уайт из отделения микроскопии и гистологии Института медицинских наук Университета Абердина, Абердин, Соединенное Королевство, за обучение работе с микроскопами и обработке с помощью SEM. Выражаю благодарность доктору Дэвиду Стэду из Абердинского центра протеомики Университета Абердина, Абердин, Соединенное Королевство, за обработку образцов белка.
Сноски
- lifescience.roche.com/en_gb/brands/universal-probe-library.html # assay-design-center
- secure.eurogentec.com/life-science.html
- geneontology.org/
Список литературы
Бернар Ф. X., Педретти Н., Росди М. и Дегерси А. (2002). Сравнение профилей экспрессии генов в однослойных культурах кератиноцитов человека, восстановленном эпидермисе и нормальной коже человека; транскрипционные эффекты лечения ретиноидами в восстановленном эпидермисе человека. Exp. Дерматол. 11, 59–74.DOI: 10.1034 / j.1600-0625.2002.110107.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Брильханте, Р. С. Н., Кордейро, Р. А., Гомес, Дж. М. Ф., Сидрим, Дж. Дж. К. и Роча, М. Ф. Г. (2006). Дерматофитоз собак, вызванный антропофильными видами: молекулярная и фенотипическая характеристика Trichophytontonsurans . J. Med. Microbiol. 55, 1583–1586. DOI: 10.1099 / jmm.0.46665-0
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Бэрден, Н., Чепмен, К., Сьюэлл, Ф., Робинсон, В. (2015). Новаторство в лучшей науке через 3rs: введение в национальный центр по замене, усовершенствованию и сокращению количества животных в исследованиях. J. Am. Доц. Лаборатория. Anim. Sci. 54, 198–208. DOI: 10.2106 / JBJS.G.01503
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Каулин К., Салвесен Г. С. и Осима Р. Г. (1997). Расщепление кератина 18 каспазой и реорганизация промежуточных филаментов во время апоптоза эпителиальных клеток. J. Cell Biol. 138, 1379–1394. DOI: 10.1083 / jcb.138.6.1379
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Дансо, М. О., Беркерс, Т., Миеремет А., Хаусил, Ф., и Баустра, Дж. К. (2015). Модель кожи человека ex vivo для изучения восстановления кожного барьера. Exp. Дерматол. 24, 48–54. DOI: 10.1111 / exd.12579
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Чу Д. Х., Хааке А. Р., Хобрук К., Лумис К.А. (2003). Строение и развитие кожи. in Дерматология Фитцпатрика в общей медицине, часть вторая . (ред.) И. М. Фридберг, А. З. Эйзен, К. Вольф, К. Ф. Остин и Л.А. Голдсмит. Нью-Йорк, Нью-Йорк: пресса MGraw-Hill.
Эмануэльссон П. и Кратц Г. (1997). Характеристика новой модели ожоговой раны in vitro. Бернс 23, 32–36. DOI: 10.1016 / S0305-4179 (96) 00073-3
CrossRef Полный текст | Google Scholar
Фэуэй, Э., Камбье, Л., Миньон, Б., Poumay, Y., и Lambert De Rouvroit, C. (2017). Моделирование дерматофитии в реконструированном эпидермисе человека: новый инструмент для изучения механизмов заражения и тестирования противогрибковых агентов. Med. Mycol. 55, 485–494. DOI: 10.1093 / MMY / myw111
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гао, Ю., Ван, X., Чен, С., Ли, С., и Лю, X. (2013). Острое нарушение кожного барьера при повторном снятии ленты: модель повреждения кожного барьера in vivo. Skin Res.Technol. 19, 162–168. DOI: 10.1111 / srt.12028
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гиллитцер Р. и Гебелер М. (2001). Хемокины в заживлении кожных ран. J. Leukoc. Биол. 69, 513–521. DOI: 10.1189 / jlb.69.4.513
CrossRef Полный текст | Google Scholar
Гребер Ф., Холейтер М., Хампель М., Хиндерер С. и Шенке-Лейланд К. (2011). Тканевая инженерия кожи — в vivo и vitro заявках. Adv. Препарат Делив. Ред. 63, 352–366. DOI: 10.1016 / j.addr.2011.01.005
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Гуджонссон, Дж. Э., Джонстон, А., Дайсон, М., Вальдимарссон, Х., и Элдер, Дж. Т. (2007). Мышиные модели псориаза. J. Investig. Дерматол. 127, 1292–1308. DOI: 10.1038 / sj.jid.5700807
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Guilloteau, K., Paris, I., Pedretti, N., Boniface, K., Juchaux, F., Huguier, V., et al. (2010). Воспаление кожи, вызванное синергическим действием IL-17A, IL-22, онкостатина M, IL-1 и TNF-повторяет некоторые особенности псориаза. J. Immunol. 184, 5263–5270. DOI: 10.4049 / jimmunol.04
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Хоми, Б., Дьё-Носжан, М. К., Визенборн, А., Массакриер, К., Пин, Дж. Дж., Олдхэм, Э. и др. (2000). Повышающая регуляция макрофагального воспалительного белка-3 / ccl20 и cc хемокинового рецептора 6 при псориазе. J. Immunol. 164, 6621–6632. DOI: 10.4049 / jimmunol.164.12.6621
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Huang, X. Z., Liang, P. P., Ma, H., Jin, S. C., Yi, J. L., Chen, Z. R., et al. (2015). Влияние культурального супернатанта, полученного из Trichophyton rubrum , выращенного в среде для ногтей, на молекулы HaCaT, связанные с врожденным иммунитетом. Подбородок. Med. J. 128, 3094–3100. DOI: 10.4103 / 0366-6999.169106
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Клещинский, К.и Фишер Т.В. (2012). Разработка краткосрочной модели культуры органов на полной толщине кожи человека in vitro в бессывороточных условиях. Arch. Дерматол. Res. 304, 579–587. DOI: 10.1007 / s00403-012-1239-z
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Кюбахер А., Зон К., Бургер-Кентишер А. и Рупп С. (2016). Модель кожи человека с добавлением иммунных клеток для изучения грибковых инфекций в программе Идентификация грибковых патогенов человека. Методы молекулярной биологии т.1508 (ред.) T. Lion 439–449. Humana Press, Нью-Йорк, штат Нью-Йорк. DOI: 10.1007 / 978-1-4939-6515-1_25
PubMed Аннотация | CrossRef Полный текст
Ли, Д. Ю., и Чо, К. Х. (2005). Влияние эпидермальных кератиноцитов и дермальных фибробластов на формирование базальной мембраны кожи в трехмерных системах культивирования. Arch. Дерматол. Res. 296, 296–302. DOI: 10.1007 / s00403-004-0529-5
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лян, П.П., Хуанг, X. Z., Yi, J. L., Chen, Z. R., Ma, H., Ye, C. X., Chen, X. Y., et al. (2016). Модель инфекции Trichophyton rubrum на основе реконструированного эпидермиса человека — Episkin ® . Подбородок. Med. J. 129, 54–58. DOI: 10.4103 / 0366-6999.172573
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ливак К. Дж. И Шмитген Т. Х. (2001). Анализ данных относительной экспрессии генов с использованием количественной ПЦР в реальном времени и метода 2-ΔΔCT. Методы 25, 402–408. DOI: 10.1006 / meth.2001.1262
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Лоссос, И.С., Червински, Д.К., Вексер, М., и Леви, Р. (2003). Оптимизация количественных параметров ОТ-ПЦР в реальном времени для исследования лимфоидных злокачественных новообразований. Лейкемия 17, 789–795. DOI: 10.1038 / sj.leu.2402881
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Марквардт Ю., Аманн, П. М., Хейсе, Р., Чая, К., Steiner, T., Merk, H. F., et al. (2015). Характеристика новой стандартизированной трехмерной модели заживления кожных ран человека с использованием непоследовательного фракционного ультраимпульсного лечения CO2-лазером. Lasers Surg. Med. 47, 257–265. DOI: 10.1002 / lsm.22341
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Méhul, B., Asselineau, D., Bernard, A. S., Leclaire, J., Régnier, M., Schmidt, R., et al. (2004). Профили экспрессии генов трех различных моделей реконструированного эпидермиса человека и классических культур кератиноцитов с использованием массивов CDNA. Arch. Дерматол. Res. 296, 145–156. DOI: 10.1007 / s00403-004-0491-2
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Накамура М., Рикимару Т., Яно Т., Мур К. Г., Пула П. Дж., Скофилд Б. Х. и др. (1990). Полнослойные эксплантаты кожи человека для тестирования токсичности химических веществ, применяемых местно. J. Investig. Дерматол. 95, 325–332. DOI: 10.1111 / 1523-1747.ep12485073
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Наканиши, К., Йошимото, Т., Цуцуи, Х., и Окамура, Х. (2001). Интерлейкин-18 регулирует ответы как Th2, так и Th3. Annu. Rev. Immunol. 19, 423–474. DOI: 10.1146 / annurev.immunol.19.1.423
CrossRef Полный текст | Google Scholar
Недошитко, Б., Соколовска-Войдыло, М., Рукеманн-Дзюрдзиньска, К., Рошкевич, Ю. и Новицки, Р. Дж. (2014). Сеть хемокинов и цитокинов в патогенезе воспалительных заболеваний кожи: атопического дерматита, псориаза и мастоцитоза кожи. Postepy Dermatol. Алергол. 31, 84–91. DOI: 10.5114 / pdia.2014.40920
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Ng, K. W., Pearton, M., Coulman, S., Anstey, A., Gateley, C., Morrissey, A., et al. (2009). Разработка модели кожи человека ex vivo для внутрикожной вакцинации: жизнеспособность тканей и поведение клеток Лангерганса. Vaccine 27, 5948–5955. DOI: 10.1016 / j.vaccine.2009.07.088
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Перес, Н.Т., Сильва, Л. Г., да Сильва, Р. С., Якоб, Т. Р., Персиноти, Ф. Г., Роча, Л. Б. и др. (2016). Модели инфекции vitro и ex vivo помогают оценить молекулярные аспекты взаимодействия Trichophyton rubrum со средой хозяина. Med. Mycol. 54, 420–427. DOI: 10.1093 / MMY / myv113
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Пирес, К. А., Круз, Н. Ф., Лобато, А. М., Соуза, П. О., Карнейро, Ф. Р., Мендес, А.М. (2014). Клинико-эпидемиологический и терапевтический профиль дерматофитии. An. Бюстгальтеры. Дерматол. 89, 259–264. DOI: 10.1590 / abd1806-4841.20142569
CrossRef Полный текст | Google Scholar
Pohin, M., Veaute, C., Garnier, J., Barrault, C., Cronier, C., Huguier, V., Favot, L., et al. (2017). Разработка новой модели реконструированного эпидермиса мыши и характеристика ее ответа на провоспалительные цитокины. J. Tissue Eng. Regen. Med. 12, e1098 – e1107 DOI: 10.1002 / термин.2442
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Пойнтнер, К., Блази, Б., Аркалис, Э., Мирастский, В., Стерфлингер, К., и Тафер, Х. (2016). Транскриптом Exophiala dermatitidis во время инфицирования модели кожи ex-vivo . Фронт. Клетка. Заразить. Microbiol. 6: 136. DOI: 10.3389 / fcimb.2016.00136
CrossRef Полный текст | Google Scholar
Пойнтнер, К., Мирастский, У., Стерфлингер, К., и Тафер, Х. (2018). Транскриптомы Изучение мутанта Exophiala Dermatitidis PKS1 на модели кожи ex vivo : важен ли меланин для инфекции? Фронт. Microbiol. 9: 1457. DOI: 10.3389 / fmicb.2018.01457
PubMed Аннотация | CrossRef Полный текст
Кейрос-Теллес, Ф., де Хуг, С., Вагнер, К. Л., Сантос, Д., Гуэдес Сальгадо, К., Висенте, В. А., Бонифаз, А., и др. (2017). Хромобластомикоз. Clin. Microbiol. Rev. 30, 233–276.DOI: 10.1128 / CMR.00032-16
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Рехави, Г., Меворах, Д., Гал, Х., Ингбер, А., Вербовецкий, И., Гивол, Д. и др. (2006). Профиль экспрессии гена UVB-индуцированного эпидермиса человека in vivo отличается от такового в культивируемых кератиноцитах. Онкоген 25, 2601–2614. DOI: 10.1038 / sj.onc.1209292
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Рендл, М., Бан, Дж., Мрасс, П., Майер, К., Ленгауэр, Б., Экхарт, Л. и др. (2002). Экспрессия каспазы-14 кератиноцитами эпидермиса регулируется ретиноидами дифференцированным образом. J. Investig. Дерматол. 119, 1150–1155. DOI: 10.1046 / j.1523-1747.2002.19532.x
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Сюй В., Чжон Хонг С., Цзя С., Чжао Ю., Галиано Р. Д. и Мустое Т. Х. (2012). Применение модели культуры кожи ex vivo человека неполной толщины в исследовании заживления кожных ран. Lab. Инвестировать. 92, 584–599. DOI: 10.1038 / labinvest.2011.184
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Zaugg, C., Monod, M., Weber, J., Harshman, K., Pradervand, S., Thomas, J., et al. (2009). Профили экспрессии генов в патогенном дерматофите человека Trichophyton rubrum во время роста на белках. Эукариот. Cell 8, 241–250. DOI: 10.1128 / EC.00208-08
PubMed Аннотация | CrossRef Полный текст | Google Scholar
Покровная система
Нажмите, чтобы просмотреть большое изображение
Продолжение сверху… повреждать. Экзокринные железы покровной системы вырабатывают пот, масло и воск для охлаждения, защиты и увлажнения поверхности кожи.Анатомия покровной системы
Эпидермис
Эпидермис — это самый поверхностный слой кожи, покрывающий почти всю поверхность тела. Эпидермис опирается на более глубокий и толстый слой дермы кожи и защищает их. Структурно эпидермис имеет толщину всего лишь около одной десятой миллиметра, но состоит из 40-50 рядов уложенных друг на друга плоских эпителиальных клеток.Эпидермис — это бессосудистая область тела, а это означает, что он не содержит крови или кровеносных сосудов. Клетки эпидермиса получают все свои питательные вещества путем диффузии жидкостей из дермы .
Эпидермис состоит из нескольких специализированных типов клеток. Почти 90% эпидермиса состоит из клеток, известных как кератиноциты. Кератиноциты развиваются из стволовых клеток в основании эпидермиса и начинают производить и накапливать протеин кератин. Кератин делает кератиноциты очень прочными, чешуйчатыми и водостойкими.Меланоциты, составляющие около 8% клеток эпидермиса, образуют второй по численности тип клеток эпидермиса. Меланоциты производят пигмент меланин, который защищает кожу от ультрафиолетового излучения и солнечных ожогов. Клетки Лангерганса являются третьими по распространенности клетками эпидермиса и составляют чуть более 1% всех клеток эпидермиса. Роль клеток Лангерганса заключается в обнаружении и борьбе с патогенами, которые пытаются проникнуть в организм через кожу. Наконец, клетки Меркеля составляют менее 1% всех клеток эпидермиса, но выполняют важную функцию — сенсорное прикосновение.Клетки Меркеля образуют диск вдоль самого глубокого края эпидермиса, где они соединяются с нервными окончаниями дермы, чтобы ощущать легкое прикосновение.
На большей части тела эпидермис состоит из 4 отдельных слоев. На ладонной поверхности рук и подошвенной поверхности стоп кожа толще, чем на остальной части тела, и имеется пятый слой эпидермиса. Самая глубокая область эпидермиса — это базальный слой, который содержит стволовые клетки, которые воспроизводятся, чтобы сформировать все другие клетки эпидермиса.Клетки базального слоя включают кубовидные кератиноциты, меланоциты и клетки Меркеля. Поверхностно по отношению к базальному слою находится шиповидный слой, в котором находятся клетки Лангерганса и множество рядов шиповидных кератиноцитов. Найденные здесь шипы представляют собой клеточные выступы, называемые десмосомами, которые образуются между кератиноцитами, чтобы удерживать их вместе и противостоять трению. Поверхностно по отношению к остистому слою находится гранулированный слой, в котором кератиноциты начинают продуцировать восковидные пластинчатые гранулы для водонепроницаемости кожи.Кератиноциты в гранулированном слое настолько удалены от дермы, что начинают умирать из-за недостатка питательных веществ. На толстой коже рук и ног есть слой кожи, расположенный поверх гранулированного слоя, известный как прозрачный слой. Люцидный слой состоит из нескольких рядов прозрачных мертвых кератиноцитов, которые защищают нижележащие слои. Самый внешний слой кожи — это роговой слой. Роговой слой состоит из множества рядов уплощенных мертвых кератиноцитов, которые защищают нижележащие слои.Мертвые кератиноциты постоянно отрываются от поверхности рогового слоя и заменяются клетками, поступающими из более глубоких слоев.
Dermis
Дерма — это глубокий слой кожи, находящийся под эпидермисом. Дерма в основном состоит из плотной соединительной ткани неправильной формы, а также нервной ткани, крови и кровеносных сосудов. Дерма намного толще эпидермиса и придает коже прочность и эластичность. Внутри дермы есть две отдельные области: сосочковый слой и ретикулярный слой.
Сосочковый слой — это поверхностный слой дермы, граничащий с эпидермисом. Сосочковый слой содержит множество пальцевидных расширений, называемых дермальными сосочками, которые выступают поверхностно по направлению к эпидермису. Дермальные сосочки увеличивают площадь поверхности дермы и содержат множество нервов и кровеносных сосудов, которые выступают к поверхности кожи. Кровь, протекающая через сосочки дермы, обеспечивает клетки эпидермиса питательными веществами и кислородом. Нервы дермальных сосочков используются для ощущения прикосновения, боли и температуры через клетки эпидермиса.
Более глубокий слой дермы, ретикулярный слой, является более толстой и жесткой частью дермы. Ретикулярный слой состоит из плотной соединительной ткани неправильной формы, которая содержит множество прочных волокон коллагена и эластина, идущих во всех направлениях, чтобы обеспечить прочность и эластичность кожи. Ретикулярный слой также содержит кровеносные сосуды, которые поддерживают клетки кожи и нервную ткань, чтобы чувствовать давление и боль в коже.
Гиподерма
Глубоко в дерме находится слой рыхлой соединительной ткани, известный как гиподерма, подкожная ткань или подкожная ткань.Гиподерма служит гибким соединением между кожей и нижележащими мышцами и костями, а также местом хранения жира. Ареолярная соединительная ткань в подкожной клетчатке содержит волокна эластина и коллагена, свободно расположенные, чтобы позволить коже растягиваться и двигаться независимо от ее нижележащих структур. Жировая жировая ткань в подкожной клетчатке хранит энергию в виде триглицеридов. Жир также помогает изолировать тело, улавливая тепло тела, вырабатываемое основными мышцами.
Волосы
Волосы — это дополнительный орган кожи, состоящий из столбов плотно упакованных мертвых кератиноцитов, обнаруженных в большинстве областей тела.Несколько безволосых частей тела включают ладонную поверхность рук, подошвенную поверхность стопы, губ , малых половых губ и головку полового члена . Волосы помогают защитить тело от ультрафиолетового излучения, предотвращая попадание солнечных лучей на кожу. Волосы также изолируют тело, задерживая теплый воздух вокруг кожи.
Волос можно разделить на 3 основные части: фолликул, корень и стержень. Волосяной фолликул — это углубление эпидермальных клеток глубоко в дерму.Стволовые клетки в фолликуле воспроизводятся, образуя кератиноциты, которые в конечном итоге образуют волосы, в то время как меланоциты производят пигмент, придающий волосам его цвет. Внутри фолликула находится корень волоса, часть волоса ниже поверхности кожи. По мере того как фолликул производит новые волосы, клетки корня поднимаются на поверхность, пока не выйдут из кожи. Стержень волоса состоит из части волоса, которая находится за пределами кожи.
Ствол и корень волоса состоят из 3 различных слоев клеток: кутикулы, коры и продолговатого мозга.Кутикула — это самый внешний слой, состоящий из кератиноцитов. Кератиноциты кутикулы наложены друг на друга, как опоясывающий лишай, так что внешний кончик каждой клетки направлен в сторону от тела. Под кутикулой находятся клетки коры, которые составляют большую часть ширины волоса. Веретенообразные и плотно упакованные клетки коры содержат пигменты, придающие цвет волосам. Самый внутренний слой волоса, мозговое вещество, присутствует не во всех волосах. Когда присутствует, продолговатый мозг обычно содержит сильно пигментированные клетки, наполненные кератином.Когда мозгового вещества нет, кора продолжается через середину волоса.
Выпадение волос происходит естественным образом у мужчин и женщин, но существуют различные методы лечения и новые способы доступа к этим медицинским услугам. Прочтите наши обзоры Hims и Hers для получения объективной информации об их планах лечения выпадения волос для мужчин и женщин соответственно. Эти компании также предлагают мощные средства по уходу за кожей, которые борются с прыщами.
Гвозди
Ногти — это дополнительные органы кожи, состоящие из листов затвердевших кератиноцитов, которые находятся на дистальных концах пальцев рук и ног. Ногти и Ногти укрепляют и защищают концы пальцев и используются для соскабливания и манипулирования небольшими предметами. Гвоздь состоит из 3 основных частей: корня, тела и свободного края. Корень ногтя — это часть ногтя, находящаяся под поверхностью кожи. Тело стержня — это видимая внешняя часть стержня. Свободный край — это дистальный конец ногтя, который вышел за пределы пальца руки или ноги.
Ногти растут из глубокого слоя эпидермальной ткани, известной как матрица ногтя, которая окружает корень ногтя. Стволовые клетки матрикса ногтя воспроизводятся с образованием кератиноцитов, которые, в свою очередь, производят белок кератина и упаковываются в жесткие слои затвердевших клеток. Листы кератиноцитов образуют твердый корень ногтя, который медленно вырастает из кожи и формирует тело ногтя, достигая поверхности кожи. Клетки корня ногтя и тела ногтя подталкиваются к дистальному концу пальца руки или ноги за счет образования новых клеток в матрице ногтя.Под телом ногтя находится слой эпидермиса и дермы, известный как ногтевое ложе. Ногтевое ложе имеет розовый цвет из-за наличия капилляров, поддерживающих клетки тела ногтя. Проксимальный конец ногтя возле корня образует беловатую форму полумесяца, известную как лунула, где через тело ногтя видно небольшое количество матрикса ногтя. Вокруг проксимального и бокового краев ногтя находится эпонихий , слой эпителия, который перекрывает и покрывает край тела ногтя.Эпонихий помогает герметизировать края ногтя, чтобы предотвратить инфицирование подлежащих тканей.
Судоносные железы
Судоносные железы — это экзокринные железы, находящиеся в дерме кожи и обычно известные как потовые железы. Есть 2 основных типа потовых желез: эккринные потовые железы и апокриновые потовые железы. Эккриновые потовые железы находятся почти в каждой области кожи и производят секрецию воды и хлорида натрия. Эккриновый пот доставляется через канал к поверхности кожи и используется для понижения температуры тела за счет испарительного охлаждения.
Апокриновые потовые железы находятся в основном в подмышечной и лобковой областях тела. Протоки апокриновых потовых желез проходят в волосяные фолликулы, так что пот, производимый этими железами, выходит из тела по поверхности стержня волоса. Апокриновые потовые железы неактивны до полового созревания, после чего они производят густую маслянистую жидкость, которую потребляют бактерии, живущие на коже. При переваривании апокринного пота бактериями возникает запах тела.
Сальные железы
Сальные железы — это экзокринные железы, находящиеся в дерме кожи, которые производят масляный секрет, известный как кожный жир.Сальные железы находятся во всех частях кожи, за исключением толстой кожи ладоней и подошв ног. Кожный жир вырабатывается сальными железами и переносится по протокам к поверхности кожи или волосяным фолликулам. Кожный жир делает кожу водонепроницаемой и повышает эластичность. Кожное сало также смазывает и защищает кутикулы волос, когда они проходят через фолликулы на внешнюю поверхность тела.
Серные железы
Церуминозные железы — это особые экзокринные железы, обнаруженные только в дерме слуховых проходов.Церуминозные железы производят восковой секрет, известный как серная пыль, для защиты слуховых проходов и смазки барабанной перепонки . Cerumen защищает уши, задерживая инородные материалы, такие как пыль и переносимые по воздуху патогены, которые попадают в ушной канал . Серума создается непрерывно и медленно выталкивает старую серную пыль наружу к наружной части слухового прохода, где она выпадает из уха или удаляется вручную.
Физиология покровной системы
Кератинизация
Кератинизация, также известная как ороговение, представляет собой процесс накопления кератина в кератиноцитах.Кератиноциты начинают свою жизнь как потомство стволовых клеток базального слоя. Молодые кератиноциты имеют кубовидную форму и почти не содержат кератинового белка. По мере размножения стволовые клетки выталкивают старые кератиноциты к поверхности кожи и в поверхностные слои эпидермиса. К тому времени, когда кератиноциты достигают шиповидного слоя, они начинают накапливать значительное количество кератина и становятся более твердыми, плоскими и водостойкими. Когда кератиноциты достигают гранулезного слоя, они становятся более плоскими и почти полностью заполняются кератином.На этом этапе клетки настолько удалены от питательных веществ, которые диффундируют из кровеносных сосудов дермы, что клетки проходят процесс апоптоза. Апоптоз — это запрограммированная смерть клетки, при которой клетка переваривает собственное ядро и органеллы, оставляя после себя только прочную, заполненную кератином оболочку. Мертвые кератиноциты, движущиеся в просвет и роговой слой, очень плоские, твердые и плотно упакованы, образуя кератиновый барьер для защиты подлежащих тканей.
Температурный гомеостаз
Будучи самым внешним органом тела, кожа способна регулировать температуру тела, контролируя взаимодействие тела с окружающей средой.В случае перехода тела в состояние гипертермии кожа способна снижать температуру тела за счет потоотделения и расширения сосудов. Пот, производимый потовыми железами, доставляет воду на поверхность тела, где она начинает испаряться. Испарение пота поглощает тепло и охлаждает поверхность тела. Расширение сосудов — это процесс, посредством которого гладкие мышцы, выстилающие кровеносные сосуды дермы, расслабляются и позволяют большему количеству крови проникать в кожу. Кровь переносит тепло по телу, отводя тепло от ядра тела и откладывая его в коже, откуда оно может излучаться из тела во внешнюю среду.
В случае перехода тела в состояние переохлаждения кожа может повышать температуру тела за счет сокращения мышц, сокращающих пили, и за счет сужения сосудов. Волосковые фолликулы имеют небольшие пучки гладкой мускулатуры, прикрепленные к их основанию, которые называются арректорными мышцами. Пили arrector образуют мурашки по коже, сокращаясь для перемещения волосяного фолликула и поднимая стержень волоса вертикально от поверхности кожи. Это движение приводит к тому, что под волосами остается больше воздуха, чтобы изолировать поверхность тела.Сужение сосудов — это процесс сокращения гладких мышц стенок кровеносных сосудов дермы для уменьшения притока крови к коже. Сужение сосудов позволяет коже охладиться, в то время как кровь остается в ядре тела, чтобы поддерживать тепло и кровообращение в жизненно важных органах.
Синтез витамина D
Витамин D, важный витамин, необходимый для усвоения кальция из пищи, вырабатывается ультрафиолетовым (УФ) светом, поражающим кожу. Слои базального слоя и шиповника эпидермиса содержат молекулу стерола, известную как 7-дегидрохолестерин.Когда ультрафиолетовый свет, присутствующий в солнечном свете или свете солярия, попадает на кожу, он проникает через внешние слои эпидермиса и поражает некоторые молекулы 7-дегидрохолестерина, превращая его в витамин D3. Витамин D3 превращается в почках в кальцитриол, активную форму витамина D. Когда наша кожа не подвергается воздействию достаточного количества солнечного света, у нас может развиться дефицит витамина D, что потенциально может привести к серьезным проблемам со здоровьем. К счастью, возможность заказать домашний тест на витамин D и проверить наш собственный уровень упрощает выявление дефицита.
Защита
Кожа обеспечивает защиту подлежащих тканей от патогенов, механических повреждений и УФ-излучения. Патогены, такие как вирусы и бактерии, не могут проникнуть в организм через неповрежденную кожу из-за того, что самые внешние слои эпидермиса содержат нескончаемый запас жестких мертвых кератиноцитов. Эта защита объясняет необходимость очистки и закрытия порезов и царапин повязками для предотвращения инфекции. Незначительные механические повреждения грубыми или острыми предметами в основном поглощаются кожей, прежде чем они могут повредить подлежащие ткани.Эпидермальные клетки постоянно воспроизводятся, чтобы быстро восстановить любые повреждения кожи. Меланоциты в эпидермисе производят пигмент меланин, который поглощает ультрафиолетовый свет, прежде чем он сможет пройти через кожу. Ультрафиолетовый свет может привести к тому, что клетки станут злокачественными, если они не будут заблокированы от попадания в организм.
Цвет кожи
Цвет кожи человека контролируется взаимодействием 3 пигментов: меланина, каротина и гемоглобина. Меланин — это коричневый или черный пигмент, вырабатываемый меланоцитами для защиты кожи от УФ-излучения.Меланин придает коже желтовато-коричневый или коричневый оттенок и обеспечивает цвет коричневых или черных волос. Производство меланина увеличивается, поскольку кожа подвергается воздействию более высоких уровней ультрафиолетового излучения, что приводит к загару кожи. Каротин — это еще один пигмент, присутствующий в коже, который придает коже желтый или оранжевый оттенок и наиболее заметен у людей с низким уровнем меланина. Гемоглобин — еще один пигмент, наиболее заметный у людей с низким содержанием меланина. Гемоглобин — это красный пигмент, содержащийся в красных кровяных тельцах, но его можно увидеть сквозь слои кожи как светло-красный или розовый цвет.Гемоглобин наиболее заметен в окраске кожи во время вазодилатации, когда капилляры дермы открыты для переноса большего количества крови на поверхность кожи.
Кожные ощущения
Кожа позволяет телу ощущать внешнюю среду, улавливая сигналы прикосновения, давления, вибрации, температуры и боли. Диски Меркеля в эпидермисе соединяются с нервными клетками дермы, чтобы определять формы и текстуры объектов, контактирующих с кожей. Тельца прикосновения — это структуры, обнаруженные в дермальных сосочках дермы, которые также обнаруживают прикосновение объектами, контактирующими с кожей.Пластинчатые тельца, обнаруженные глубоко в дерме, ощущают давление и вибрацию кожи. По всей дерме есть множество свободных нервных окончаний, которые представляют собой просто нейроны, дендриты которых распространены по всей дерме. Свободные нервные окончания могут быть чувствительны к боли, теплу или холоду. Плотность этих сенсорных рецепторов в коже варьируется по всему телу, в результате чего некоторые области тела более чувствительны к прикосновению, температуре или боли, чем другие области.
Экскреция
Помимо выделения пота для охлаждения тела, эккринные потовые железы кожи также выводят из организма продукты жизнедеятельности.Пот, вырабатываемый эккринными потовыми железами, обычно содержит в основном воду с большим количеством электролитов и некоторыми другими химическими веществами в следовых количествах. Наиболее распространенными электролитами, обнаруженными в поте, являются натрий и хлорид, но также могут выводиться ионы калия, кальция и магния. Когда эти электролиты достигают высокого уровня в крови, их присутствие в поте также увеличивается, что помогает уменьшить их присутствие в организме. Помимо электролитов, пот содержит и помогает выводить небольшое количество продуктов метаболизма, таких как молочная кислота, мочевина, мочевая кислота и аммиак.Наконец, эккринные потовые железы могут помочь вывести алкоголь из организма человека, употребляющего алкогольные напитки. Алкоголь вызывает расширение сосудов в дерме, что приводит к усилению потоотделения по мере того, как больше крови достигает потовых желез. Алкоголь в крови поглощается клетками потовых желез, в результате чего он выводится вместе с другими компонентами пота.
клеток кожи — Science NetLinks
Назначение
Для развития понимания различных типов клеток, из которых состоит кожа человека, и того, как разные клетки кожи связаны друг с другом.
Контекст
Этот урок является частью проекта Skin Deep, который исследует науку, лежащую в основе кожи. Skin Deep разработана AAAS и финансируется Neutrogena. Для получения дополнительных уроков, занятий и интерактивных материалов, которые ближе познакомятся с наукой, лежащей в основе кожи, обязательно посетите страницу проекта Skin Deep.
На этом уроке учащиеся используют онлайн-ресурсы и проводят лабораторный эксперимент, чтобы узнать о различных типах клеток, составляющих кожу, и о том, как разные клетки кожи связаны друг с другом.
Для учащихся на этом уровне важно начать понимать, что все самовоспроизводящиеся формы жизни состоят из клеток, от одноклеточных бактерий до слонов с их триллионами клеток. Невооруженным глазом можно увидеть несколько гигантских клеток, таких как куриные яйца. Именно на клеточном уровне выполняются многие из основных функций организмов: синтез белка, извлечение энергии из питательных веществ и т. Д. Все живые клетки имеют похожие типы сложных молекул, которые участвуют в этих основных жизненных действиях.Этот урок начинает показывать, что в дополнение к основным клеточным функциям, общим для всех клеток, большинство клеток в многоклеточных организмах выполняют некоторые особые функции, которых нет у других. ( Наука для всех американцев , стр. 80-82.)
Что касается клеток, многие студенты могут полагать, что организмы содержат клетки, а не состоят из клеток. Представление о большом количестве клеток также может быть проблемой. Крупные организмы состоят примерно из триллиона клеток. Однако этот большой номер может быть трудным для понимания.Чтобы сделать его менее абстрактным, студенты могут лучше понять миллион миллионов, чем триллион.
Студентам может быть еще труднее понять, что клетки являются основными единицами, в которых происходят жизненные процессы. По этим причинам рассказ о клетках должен быть простым. Однако понимание основных клеточных процессов является необходимым условием, которое следует изучить до этого урока.
Студенты будут использовать микроскопы на этом уроке, поэтому они должны знать, как пользоваться микроскопом, и знать его части.Они также должны знать надлежащие лабораторные процедуры и правильную утилизацию лабораторных материалов, в том числе острых предметных стекол и химикатов. Обладая базовым «чувством увеличения», учащиеся могут использовать микрофотографии для расширения своих наблюдений за клетками, которые могут начать показывать учащимся, что клетки являются фундаментальными строительными блоками их собственных тел, а также других живых существ. ( Benchmarks for Science Literacy , pp. 110-112). Студентам также может быть легче понять, что клетка является основной структурной единицей (которую они могут наблюдать), чем то, что клетка является основной единицей функции ( что должно быть выведено из экспериментов).( Контрольные показатели научной грамотности , стр. 342)
Планирование вперед
Примечание. Для получения предварительных знаний учащиеся должны заполнить Ячейки 2: Ячейка как система, охватывающая базовую структуру ячеек и системы, прежде чем начать этот урок.
Мотивация
Чтобы заинтересовать учащихся и сфокусировать их на теме, напишите на доске слово «кожа» и предложите им провести мозговой штурм по словам, концепциям или процессам, которые приходят в голову.Запишите все ответы (например, волосы, морщины, пот) и попросите класс обсудить список.
Затем просмотрите, что студенты уже знают о коже и клетках в целом, задав такие вопросы:
- Что кожа делает для тела?
- (Кожа помогает защитить тело от травм; обеспечивает защиту от вредных веществ и организмов, таких как бактерии и паразиты; помогает удерживать влагу в теле; помогает поддерживать постоянную температуру тела; и имеет нервные окончания, которые помогают вам чувствовать ощущения.)
- Что было бы, если бы у вас не было кожи?
- (Ответы могут быть разными, но побудите студентов понять, что кожа — наша первая линия защиты от вреда.)
- Из чего состоит кожа?
- (состоит из клеток кожи.)
- Что делают клетки?
- (Клетки выполняют три основные функции: клетки растут за счет функционирования клеточного метаболизма; клетки создают новые клетки посредством деления клеток; и клетки способны синтезировать новые белки, которые необходимы для модуляции и поддержания клеточной активности.)
- Все ли ячейки одинаковы?
- (№. Клетки различаются в зависимости от выполняемой ими работы. Специализированные клетки [например, мышечные клетки, нервные клетки] выполняют специализированные функции [например, вырабатывают гормоны, переносят кислород].)
- Работают ли клетки по отдельности или вместе с другими клетками?
- (Клетки обычно работают с другими клетками для выполнения определенной функции. Специализированные клетки группируются и совместно работают, чтобы сформировать ткани, такие как мышцы или жир.Они работают вместе как система.)
Разработка
В этой части урока учащиеся узнают больше о различных типах клеток, из которых состоит кожа человека.
Разделите класс на пары. Попросите каждую пару использовать свою студенческую ведомость «Клетки кожи», чтобы посетить и прочитать двухстраничное введение в клетки в «Клетки в вашем теле». Когда они закончат чтение, проверьте их понимание с помощью таких вопросов (студенты могут написать свои ответы на эти вопросы в студенческом листе Skin Cells):
- Из чего состоит все живое?
- (Они состоят из ячеек.)
- Как называются организмы с более чем одной клеткой?
- (Их называют многоклеточными организмами.)
- Сколько различных типов клеток в организме человека?
- (Существует около 200 различных типов ячеек.)
- Что есть у всех клеток и что они делают?
- (У них есть мембрана, которая удерживает клетку вместе; ядро, которое содержит информацию, которая позволяет клетке воспроизводиться; и митохондрия, где пища и кислород объединяются для производства энергии.)
- Что создают группы ячеек разных типов?
- (Они составляют органы вашего тела.)
- Что представляет собой группа различных органов, работающих вместе для выполнения работы?
- Могут ли наши клетки умереть? Если да, то заменяются ли они?
- (Да, наши клетки могут умереть, и они будут заменены. Существует широкий разброс в скорости любой смерти и регенерации, известной как оборот клеток.Это изменение зависит от типа клеток и органа. Например, люди теряют тысячи клеток кожи в день, и они быстро заменяются. Клетки мозга, однако, обычно не регенерируют.)
Также проведите обзор частей ячеек. Убедитесь, что учащиеся помнят и понимают расположение и функцию ядра, цитоплазмы и клеточной мембраны (как описано в предварительном уроке «Клетки 2: Клетка как система»).
Перед тем, как учащиеся выполнят следующее задание, предоставьте им готовые слайды с изображением клеток человеческих органов, чтобы они могли наблюдать их под микроскопом.Вы можете получить эти слайды из Microscope Slide Kit: Histology, Human Organs, из Microscope World, или вы можете показать студентам изображения различных клеток органов на Подготовленных слайдах по анатомии и бактериологии человека. Чтобы помочь студентам понять, что они видят, покажите им увеличенное изображение сквамозных эпителиальных клеток щеки из Интернет-учебника естественной истории Уэйна Word. Объясните: видимые здесь структуры нельзя увидеть без увеличения. Укажите на отмеченные части изображения и помогите учащимся понять, что они будут искать похожие структуры в своих образцах.
Затем попросите учащихся использовать свои лабораторные листы «Наблюдение за клетками кожи», чтобы посмотреть на свои собственные клетки кожи под микроскопом. Им следует работать со своими партнерами, чтобы собрать и изучить некоторые из клеток своей кожи. Студенты должны иметь опыт правильного использования микроскопов и подготовки предметных стекол. Они также должны знать надлежащие лабораторные процедуры безопасности и правильную утилизацию лабораторных материалов, в том числе острых предметных стекол и химикатов. Спросите студентов, какова функция метиленового синего.Попросите их взглянуть на свои клетки без метиленового синего, чтобы они увидели преимущества использования красителя.
Примечание. Убедитесь, что ваши ученики знают, что метиленовый синий — жизненно важное пятно: он окрашивает почти все, включая кожу и одежду. Студенты должны носить химически стойкие перчатки и избегать контакта с глазами и кожей. Они должны носить защитные очки при работе с химикатами, нагревателем или стеклянной посудой в лаборатории.
После того, как студенты завершили лабораторную работу, задайте следующие вопросы:
- Что вы заметили в клетках кожи до использования метиленового синего?
- Как метиленовый синий помогает вам более четко видеть клетки кожи?
- Какие части клетки вы видите в микроскоп и при каком увеличении вы их видите?
- Сравните этот слайд с клетками кожи со слайдами клеток человеческого органа, которые вы видели ранее.Они выглядят одинаково? Все ли клетки разных органов выглядят одинаково? Почему или почему нет?
- Вы видите одни и те же структуры среди всех различных клеток органов?
- Видите ли вы разные структуры между клетками разных органов?
(Ответы могут быть разными. Поощряйте студентов объяснять свои ответы.)
Оценка
Разделите класс на три группы, представляющие эти слои или структуры кожи:
- эпидермис
- дерма и гиподерма
- потовые и сальные железы; волосы и ногти; и цвет кожи.
Раздайте студенческий лист «Под кожей» и просмотрите его вместе с классом. Попросите учащихся в каждой группе прочитать части своего слоя или структуры и просмотреть свои заметки, чтобы убедиться, что они понимают каждую из них. Затем проинструктируйте членов группы выбрать одну маркированную часть, чтобы «стать», убедившись, что все части представлены. Каждый студент должен изучить свою часть и быть готовым объяснить, как она функционирует и взаимодействует с другими клетками кожи.
В дополнение к объяснению того, как его или ее часть функционирует и взаимодействует с другими клетками кожи, студент должен рассмотреть вопрос: что может произойти, если определенный тип клеток кожи отсутствует или поврежден? Студенты также могут найти информацию о кожных заболеваниях, которые могут повлиять на изучаемую часть кожи.
Когда все участники закончат свое исследование, попросите их представить информацию классу. Они могут сделать это в виде постерной презентации, PowerPoint и т. Д.
Расширения
Вы можете расширить представления о оболочке и системах в этом уроке, проведя студентов через эти уроки Science NetLinks:
Студенты могут весело провести время, узнав больше о структуре и функциях клеток, посетив CELLS Alive !, который содержит подробные диаграммы клеток, трехмерные модели и анимированные камеры клеток.
Cell Project от Access Excellence позволяет учащимся расширить свои знания о клетках, включая построение модели гигантской клетки.